Laws of learning in the spatial domain
We have illustrated several examples of general laws of learning in
the spatial domain in pigeons. These included overshadowing, spatial generalization,
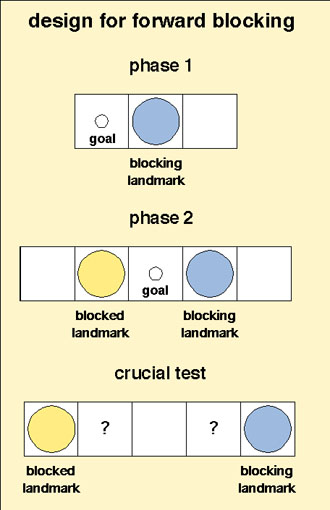
and peak shift. Other general laws could be pursued. Blocking is one example, demonstrated in rats so far in the spatial
domain (Roberts & Pearce, 1999; Rodrigo, Chamizo, McLaren, & Mackintosh,
1997). One paradigm for a touch-screen or 'arena' experiment is illustrated
here:
Odometry
How do birds estimate the distance that they have flown or walked?
Honeybees have been tested in a tunnel like the one pictured below
(Srinivasan, Zhang, & Bidwell, 1997).  In these experiments, the top of
the tunnel was covered with perspex, and the walls were lined with texture
of some kind (regular stripes or random patterns of black and white squares).
A feeder was placed some distance into the tunnel, and the bees were trained
to fly into the tunnel to obtain sugar water. After sufficient training,
the reward was removed for occasional tests. On tests, various manipulations
were made using the transformational approach.
The honeybees flew into the tunnel on tests, and stopped around the training
distance, and turned around. They often then turned back and forth a number
of times. Manipulations showed that they did not count stripes or use the
size of the entrance or back wall. Adding wind was also ineffective in changing
distance estimates, ruling against the use of energy expenditure as an estimate
of distance. The bees did use the texture on the walls, however, as removing
such texture led to spectacular nonperformance: they flew back and forth
from one end to the other! Changing the width of the tunnel altered the
distance flown. This is consistent with the use of visual flow integrated
over time as a means of distance estimation. The narrower the tunnel, the
faster the visual flow, and the less distance the insects have to fly before
the target odometric reading is reached; this is precisely the pattern found.
Other experiments (Srinivasan, Zhang, Lehrer, & Collett, 1996) showed
that the bees flew so as to maintain a constant retinal velocity. This means
that they slowed down as the tunnel narrowed, and sped up as it widened.
One purpose this can serve is landing. The mechanism ensures that the insect
slows down to zero as it approaches the landing surface.
In these experiments, the top of
the tunnel was covered with perspex, and the walls were lined with texture
of some kind (regular stripes or random patterns of black and white squares).
A feeder was placed some distance into the tunnel, and the bees were trained
to fly into the tunnel to obtain sugar water. After sufficient training,
the reward was removed for occasional tests. On tests, various manipulations
were made using the transformational approach.
The honeybees flew into the tunnel on tests, and stopped around the training
distance, and turned around. They often then turned back and forth a number
of times. Manipulations showed that they did not count stripes or use the
size of the entrance or back wall. Adding wind was also ineffective in changing
distance estimates, ruling against the use of energy expenditure as an estimate
of distance. The bees did use the texture on the walls, however, as removing
such texture led to spectacular nonperformance: they flew back and forth
from one end to the other! Changing the width of the tunnel altered the
distance flown. This is consistent with the use of visual flow integrated
over time as a means of distance estimation. The narrower the tunnel, the
faster the visual flow, and the less distance the insects have to fly before
the target odometric reading is reached; this is precisely the pattern found.
Other experiments (Srinivasan, Zhang, Lehrer, & Collett, 1996) showed
that the bees flew so as to maintain a constant retinal velocity. This means
that they slowed down as the tunnel narrowed, and sped up as it widened.
One purpose this can serve is landing. The mechanism ensures that the insect
slows down to zero as it approaches the landing surface.
We think that pigeons are likely to use retinal flow in distance estimation
as well. Functionally, for a flying animal retinal flow is the most reliable
source for estimating distance flown. Wind conditions make energy expenditure
an unreliable indicator of distance flown. Neurophysiologically, detectors
of retinal flow have been found in the pigeon's brain (Frost & Sun, 1997). The behavioral research, however, remains to be
done.
Averaging vectors used in trajectories
In measuring vectors from landmarks, pigeons
compute distance and direction independently. How do birds average vectors when the vectors are trajectories to be
flown or walked from a starting point? Research on trajectories of bees (Collett, Baron, & Sellen, 1996)
shows that the insects average entire vectors and do not average distance
and direction separately. These experiments on trajectories in honeybees were
conducted using enclosed arenas. The bee's task was to enter the arena,
and then strike a particular trajectory, for example 50 cm at 60 degrees
to the right. They then dropped through a hole on the floor to another chamber
below, in which they had to strike a different trajectory. Stripes on the
back wall, oriented at different orientations, provided discriminative stimulus.
On crucial tests, stripes at an intermediate orientation were presented
during their first flight. The logic of experimentation followed that presented
in the section on vector vs. independent
averaging. Results favored vector averaging in that they flew at an
intermediate direction and adjusted distance flown according to vector averaging.
It makes functional sense to average entire vectors describing trajectories.
It makes no sense to average the components of trajectories, distance and
direction, separately. The average of 5 m N and 5 m E from one's starting
point is not 5 m NE (result of independent averaging of direction and distance
components). Entire vectors must be averaged to get the right answer. To
path integrate, an animal must update both direction and distance simultaneously
at all times. Do birds also average whole trajectories?