III. Motion Discrimination
This section summarizes some of the main findings
about pigeons' ability to discriminate stimuli that involve some kind of
movement (see also Lea & Dittrich, 1999). But since the ability to
detect and to discriminate motion information depends on structural characteristics
of the visual system, we will give first some basic details about birds',
and in particular pigeons', eyes and the visual pathways to which they
connect. A fuller treatment can be found in Husband & Shimizu's chapter
of this volume. We also need to reflect a little on the characteristics
of the apparatus that is commonly used to display moving stimuli, i.e.
video and computer screens of various sorts. These are discussed more fully
by D'Eath (1998) and Lea and Dittrich (1999).
Differences
between bird and mammal visual systems
Butler and Hodos (1996) have reviewed the evolution
and adaptation of neuroanatomical structures across the vertebrates, and
from their highly informative book one can get an idea of the divergences
between avian and mammalian visual systems. But of course we cannot really
talk about a "typical" avian visual system. Like mammals, birds differ
greatly in many aspects of their visual function, for example in the size
and direction of their visual fields (Sillman, 1973; Martin, 1999), in
their visual acuity, and in the dependence of visual acuity on light level
are also wide (Martin, 1993). All the same, there are some consistent differences
between the two groups, and some of these may have implications for movement
perception. For example, because birds generally have flatter eyeballs
than mammals, they have severe limitations on swiveling their eyeballs,
so they rely on head movements instead to bring the important parts of
the scene onto areas of its retina with high photoreceptor density. These
head movements will necessarily also give motion parallax information,
though of course it does not follow that the bird makes use of it. We suggest
that in most non-flying situations the visual stability of the environment
is not disrupted by birds' head movements leading to retinal change but,
on the contrary, that head movements are compulsory to maintain visual
stability. What appears at first glance paradoxically is a hypothesis that
is firmly grounded on our notion of 'motion integration centers' (see above)
and the structure of the birds' visual system. Unlike mammals, about half
of the birds species that have been studied have more than one fovea on
each retina. These dense cell areas are distributed in very different places
in different bird species, leading to very different orienting head movements.
Moreover, the cell density in extrafoveal areas of the retina is generally
higher than in humans, as is the relative number of cones (see Butler &
Hodos, 1996).
These structural differences and variations
do not make the study of birds' vision uninteresting to those whose primary
interest is in human vision. On the contrary, a comparative approach offers
an essential perspective. So long as it is based on biological principles,
and not on mere botanizing, it is the one approach that can enable us to
understand the evolutionary and ecological demands on visual perception
and cognition (e.g. Kamil, 1988; Cook, 1993; Dittrich, Gilbert, Green,
McGregor & Grewcock, 1993; Shettleworth, 1993; Lea & Dittrich,
1999).
Distinctive
features of pigeon vision
Pigeons have a very wide visual field - of around
340°, compared to the 180°of humans. This is in common with most
birds (though not all - consider owls) and many mammals (e.g. rats or rabbits).
Given the high general cone density of birds, this means that in principle
at least a pigeon can track an externally moving object without moving
its head or eyes over much greater displacements than a human can. Pigeons
are among the birds with a dual fovea (e.g. Hayes, Hodos, Holden,&
Low, 1987; Husband & Shimizu, this volume (section IV)), and the two
visual systems, frontal and lateral, seem to have rather different properties
(e.g. Nye, 1973; Friedman, 1975; Martinoya et al., 1984); this means that
an object that moves between them could pose problems for the bird's perception,
unless there are mechanisms to compensate for the changes in the stimulation
it will cause.
Features
of the motion displays
Video and computer screens have a number of
well-known problems as stimulus generators for avian visual perception.
They have limited resolution; they depend on apparent movement; their color
generation is attuned to the human eye, not to the different photopigment
set of the pigeon or any other species of bird, and in particular they
emit no ultra-violet, which is involved in birds' color vision; they may
flicker at rates that are perceptible to birds but not to the human experimenter;
and of course they are inherently two-dimensional. We have explored these
problems in detail elsewhere (Lea & Dittrich, 1999), and here simply
state the conclusion we drew in that paper: that the video display is,
if not a perfect tool, a highly acceptable tool for investigating birds'
perception and cognition of visual movement. Birds may have particular
difficulties in extracting from video displays the kind of fine detail
required for discriminating between individual conspecifics, but many other
kinds of discrimination can be made successfully. So in this chapter, we
shall proceed without further comment to use such experiments to tell us
what kinds of motion discriminations pigeons and other birds can make.
Is it moving
or not and does it matter?
Two fundamental questions are whether birds
can discriminate moving from static images, and whether they can generalize
between moving and static versions of the same object. The first of these
questions has been addressed by Siegel (1970), who showed that pigeons
could discriminate simple static shapes from the same shapes in apparent
movement at speeds of between 4 and 64 Hz. In further experiments, pigeons
were able to discriminate between non-moving vertical lines, pulsating
stimuli and those in apparent or real movement (Siegel, 1971). These findings
can be seen as initial evidence that the positional change alone seems
a salient feature used for discriminating simple stimuli. Hodos, Smith
& Bonbright (1975) and Mulvanny (1978) investigated thresholds for
movement detection, and found them to range from 4.1 to 6.1°/sec. However,
these values are for frontal-viewing conditions and it can be expected
that under lateral viewing conditions (e.g., optic flow) threshold levels
may vary, as the lateral field seems to be adapted to faster moving stimuli.
Threshold differences between different groups of birds depending on their
ecology can also be expected. Dittrich and Lea (1993) extended the study
of movement detection to much more complex movements, by training pigeons
to discriminate between video scenes (of other pigeons performing characteristic
responses) and still frames from the same scenes.
Interestingly, there was a strong feature-positive
effect in this experiment, with movement-positive discriminations being
much easier to demonstrate than movement-negative discriminations with
the same stimuli; this suggests, in line with the ecological data mentioned
above, that movement is highly salient for pigeons just as it is for humans.
The converse question, of generalization from static to moving versions
of the same stimuli, has also been addressed using category discrimination
techniques. In an unpublished experiment, Watanabe, Lea, Ryan, and Ghosh
trained pigeons to discriminate between a small set of pictures of birds
and and a similar set of pictures of trees. For example, one of the
tree pictures showed an oak and one of the bird pictures showed a
pigeon (other examples
included a gull, duck and hawk, and other trees used included a fir,
Japanese cherry, and scholar tree). After training, tests were given
in which the original training stimuli were presented in either smooth
or staccato motion: the birds showed clear generalization of the discrimination
to these new versions of the stimuli, showing that movement did not render
the stimuli unrecognizable.
To see examples of the stimuli used by Watanabe, Lea, Ryan and
Ghosh, click on each of the above image to see them in smooth motion.
To see the staccato motion conditions click here (example of tree,
example of bird).
Discriminating
between different kinds of movement
Simply recognizing whether or not something
is moving is not a very interesting, however. It is more interesting to
know whether birds can discriminate between different kinds of movement.
Once again we have evidence from experiments both with simple and with
more complex stimuli. As regards simple shapes, Siegel (1971) successfully
trained pigeons to recognize the horizontal movements of approximately
three vertical lines projected onto the response key. In a generalization
test in which the horizontal movements were altered to vertical movements
in steps of 22.5º pigeons showed a relatively steep generalization
gradient as the direction of movement departed from the original horizontal
movement displays. Furthermore, he also claimed perfect transfer between
real and apparent movements in test trials. Pisacreta (1982) trained birds
to track shapes moving, in a staccato fashion, at characteristic rates
across a screen; changing the movement rate led to some generalization
decrement, though not as much as changing color or form.
A more complex case was studied by Emmerton
(1986), in a rather more sophisticated investigation. She showed that when
monochromatic stimuli were displayed on a fast oscilloscope screen, pigeons
could be trained successfully to discriminate movement patterns. With the
technology she used, there should be no uncontrolled problems with flicker,
and color was irrelevant, so many of the difficulties of using video displays
were overcome. The stimuli used were Lissajous
figures, which are produced by moving a dot on an oscilloscope screen
with independent sinusoidal motions in the vertical and horizontal dimensions;
if the ratio of the frequencies is slightly different from an integer,
the pattern generated changes constantly. If the dot moves quickly,
an outline shape is seen, though its form changes. If the dot moves
slowly, then a moving dot is seen, tracing the outline. If the ratio
of the frequencies of movement in the X and Y dimensions are changed, the
shape generated by the moving dot changes.
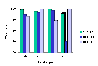 Click
here to see the results from Experiment 3 of Emmerton (1986), in which
pigeons discriminated between Lissajous figures generated by X:Y frequency
ratios of approximately 2:1 and 1:1. Of the four test types, "train"
used the same stimuli and procedure as in training; "test" used the same
stimuli but the reward contingencies were suspended; "nonrotate" again
used the same stimuli as in training, while "rotate" used stimuli in which
either the figure being traced was rotated through 90 degrees, or the direction
of rotation of the dot was reversed.
Click
here to see the results from Experiment 3 of Emmerton (1986), in which
pigeons discriminated between Lissajous figures generated by X:Y frequency
ratios of approximately 2:1 and 1:1. Of the four test types, "train"
used the same stimuli and procedure as in training; "test" used the same
stimuli but the reward contingencies were suspended; "nonrotate" again
used the same stimuli as in training, while "rotate" used stimuli in which
either the figure being traced was rotated through 90 degrees, or the direction
of rotation of the dot was reversed.
Interestingly, Emmerton showed that pigeons
generalized to new views of outline figures but failed to generalize to
views of dot-defined figures. Directional invariance seemed to hold only
for a dot moving in a circle. She interpreted these test findings as evidence
for stimulus-dependent generalization in pigeons. The only study which
seems to have continued Emmerton's theme seems Cook and Katz's (1999) study
on the perception of dynamic object properties. They trained four pigeons
to discriminate computer-generated 3-D projections of cube and pyramidal
objects. Generalization tests involved static and rotating presentations
or modifications of these stimuli (for further details see Cook, this volume).
It is interesting in the context of this chapter that they reported a dynamic
superiority effect, i.e. dynamic conditions were always discriminated better
than the static conditions (Cook & Katz, 1999, Fig.5). They interpreted
this findus diagram and diagram of Emmerton results as indication that
the objects' motion contributed information about their structure that
was not obtainable from static presentations. These findings seem to strengthen
the view that simple object motion can play a vital role in complex object
recognition tasks.
Although most of Emmerton's (1986) patterns
were complex in themselves, the particular figures used as positive and
negative stimuli were consistent from trial to trial, so the experiment
was not a category discrimination in the way that so many movement discrimination
tasks necessarily are. This difference can be highlighted when we compare
pigeons' discrimination performance between artificial Lissajous figures
and natural images. Pigeons successfully trained to discriminate between
static and motion images of conspecifics showed remarkable response differences
in generalizing between various types of movement (Dittrich & Lea,
1993, Figure 6). The authors reported a perfect negative correlation between
the responses of the motion+ and the static+ group during generalization
as well as a significant interaction between training group and type of
motion displayed. Furthermore, discrimination was shown to be independent
of size, perspective or viewing angle, brightness, and color. Both aspects
of the findings were interpreted as evidence that pigeons not only can
discriminate natural images on the basis of motion cues alone they also
seem to form a concept of motion spontaneously. The next question to ask
seemed at this stage whether pigeons can discriminate between exemplars
of a motion concept and on what basis such a motion concept would be formed.
Dittrich, Lea, Barrett and Gurr (1998) trained pigeons in a true concept
discrimination based on movement types. In their Experiments 1 and 2, they
demonstrated that pigeons could be trained to distinguish moving images
of conspecifics from one another on the basis of the movement's category
membership such as pecking or walking. Examples of the two ways of presenting
the stimuli and the two types of movement presented are chosen from Dittrich
et al (1998; click on images to see videos):
|
Category Membership
|
Full Detail
|
Point Light
|
| Pigeon
Pecking |

|

|
| Pigeon
Walking |

|

|
Several different scenes were used for each
category, so presumably several different cues would be needed to make
a reliable discrimination between the two sets (cf. Lea, 1984). Furthermore,
in their Experiment 1 they used the standard 'pseudoconcept' control procedure:
there was an additional group of birds for whom the positive and negative
sets cut across the natural classification of the stimuli. The birds trained
in this 'perverse pseudoconcept' task uniformly failed to discriminate,
so we can assume that movements that belong to the same category to human
eyes are also similar to one another from the pigeon's point of view. Lea
et al. (in prep.) successfully trained bantam hens to discriminate the
same set of video images of pigeons pecking and walking as Dittrich et
al (1998) had used . This result (see section IV) shows, not just that
the capacity to make discriminations between categories of movements is
not confined to pigeons (which would have been very surprising), but also
that it is not confined to images of conspecifics, which might be supposed
to have some special status. In fact, as in experiments using color slide
stimuli (Ryan & Lea, 1994), the chickens learned to discriminate pigeon
stimuli rather faster than pigeons did.
Optic flow
As animals move around, most of the time they
generate an informative pattern of optical changes. The optic array of
natural surfaces, whether ground, sky, or water, registers a pattern of
continuous changes with the animal's movement. This is called optic flow.
For example, when moving forward or in parallel to or directly approaching
a surface, there is a continuous flow of retinal projections relative to
the animal's viewpoint. The information in this optic flow pattern is a
reliable source for checking the relative velocity and the direction of
movement. There is good evidence that birds possess the neurological apparatus
to respond to optic flow variations (see, e.g., Wang & Frost 1992;
Wylie, Linkenhoker,& Lau, 1997; Wylie, Bischof,& Frost, 1998),
and some birds at least have a clear ecological need to do so (see, e.g.,
Lee & Reddish, 1981, who analyze the case of plunge-diving in gannets).
Recently, Sun and Frost (1998) tested the response of neurons to the image
expansion of objects approaching on a direct collision course with the
bird. Three types of looming-sensitive neurons were described in the nucleus
rotundus. Remarkably, none of these neurons seem to respond to a simulated
approach of a stationary object. The authors suggest that these neurons
provide precise information for the time-to-contact variable or the computation
of tau. Similarly, behavioral experiments to link these two lines of research
are obviously called for.
Animals seem to have at least two methods for
timing their reactions appropriately when embarked on a collision course
either with an object or surface. First, they could estimate the time remaining
before collision, or "time to contact": this could be derived from perception
of the distance and speed of the movement of the object. However, neither
of those would be directly available. Second, as Lee (1976) suggested,
the ratio between retinal image size at a given instant and the rate of
expansion of the image could be used. Lee gave this ratio the name tau.
The ratio seems sufficient as long as the image size is not too large and
the velocity of the approaching event relative to the animal is kept constant.
Its value is independent of the speed and size of the object. It has also
been reported that the landing responses of pigeons correlate well with
tau (Lee, Davies, Green & van der Weel, 1993). Lee's time-to-collision
ratio (tau) became a paradigm example of Gibson's ecological approach because
tau requires no complex internal calculations in the brain and seems readily
available from the flow field over the retina of each eye. So far, electrophysiological
recordings (see above) from single neurons in the pigeon provide the best
evidence for Lee's time-to-collision concept. Recently, it has been demonstrated
that the information used in judging time to collision events seems of
many origins (of which tau is just one) depending also, for example, on
the task and stimulus condition (see e.g. Tresilian 1993; Wann 1996). Following
our arguments on motion integrators one could say that not the detection
of a single cue, namely tau, seems sufficient but the interactive processing
of various motion parameters depending on the behavioral demands. More
behavioral studies in this area are urgently needed to understand the role
of motion variables in the determination of birds' reactions when approaching
objects or surfaces.
Next section: Motion
Recognition


