I. Introduction
The business of foraging requires an well-coordinated
group of sensory, motor, and cognitive skills. In order to complete an
act of feeding, birds must encounter, identify, select, capture, and consume
their prey. All of these behaviors depend, of course, on the needs and
the sensory equipment of the particular forager. For example, the owl,
which hunts in dim light, uses auditory cues to locate rodents. The kiwi, a crespular flightless species pictured to the right, locates underground worms
through its nostrils, which are well-placed at the tip of its long beak.
Cormorants, which dive for food, possesses a powerful visual accommodation
system that permits it to adjust its eyes to the differing optical requirements
of air and water. Nonetheless, these and other wild species appear to share
efficient food-seeking strategies that balance their nutritive intake against
energy expenditure and other costs associated foraging activities. In this
chapter I focus on the ways in which ground-feeding birds process visual
information in order to accomplish early stages of the foraging act, specifically
prey identification and choice.
flightless species pictured to the right, locates underground worms
through its nostrils, which are well-placed at the tip of its long beak.
Cormorants, which dive for food, possesses a powerful visual accommodation
system that permits it to adjust its eyes to the differing optical requirements
of air and water. Nonetheless, these and other wild species appear to share
efficient food-seeking strategies that balance their nutritive intake against
energy expenditure and other costs associated foraging activities. In this
chapter I focus on the ways in which ground-feeding birds process visual
information in order to accomplish early stages of the foraging act, specifically
prey identification and choice.
Foraging studies have considered a wide variety
of avian species, such as great tits. Much of this work has been observational, conducted
by researchers during visits to the bird’s natural habitat. Other
experiments have taken place in laboratories, and, in these cases, the
variety of species is relatively restricted for ethical and practical reasons.
Work in our laboratory, like so much else described in this book, has used
pigeons. To the observer in a city park this bird’s task may not seem particularly
challenging, but in less-friendly habitats it demands specialized cognitive
abilities to direct an efficient strategy. For visual information
processing, the needed skills fall roughly into two classes: Those that
depend mainly on visual perception and those that rely more specifically
on memory.
been observational, conducted
by researchers during visits to the bird’s natural habitat. Other
experiments have taken place in laboratories, and, in these cases, the
variety of species is relatively restricted for ethical and practical reasons.
Work in our laboratory, like so much else described in this book, has used
pigeons. To the observer in a city park this bird’s task may not seem particularly
challenging, but in less-friendly habitats it demands specialized cognitive
abilities to direct an efficient strategy. For visual information
processing, the needed skills fall roughly into two classes: Those that
depend mainly on visual perception and those that rely more specifically
on memory.
Perceptual Requirements

In
natural settings preferred foods are often difficult to detect. Indeed,
crypsis is a common adaptation on the part of the prey itself, which usually
are more likely to survive when hidden. For instance, in the patch of Florida
beach being explored by the sandpipers to the right there are pieces of seafood,
but they are concealed among pebbles,
sand, and shells. In another setting, moths often rest
on trees having bark of an appearance similar to their own (Pietrewicz
and Kamil, 1979). The prey sought by ground-feeding birds are varied in
appearance and rest on a camouflaging background composed of leaf
litter, stones, plants of various forms and colors, and other debris. In
return, the predator also adapts, benefiting from a perceptual system that
is specialized according to its specific foraging needs and the habits
of the prey. Pigeons are aided by a visual system that resolves fine visual
detail and permits subtle color discriminations.
 Visual Structure.
The
pigeon's visual system has certain features that facilitate the foraging task.
The eyes themselves are very large, filling much of the head’s interior.
While they are positioned laterally, the pupils are directed slightly forward,
providing a binocular field of view. A rich retinal structure
includes a large proportion of cones, the receptor specialized for daylight
vision. Cone pigments also provide the basis for hue discrimination. In
pigeons, as in many birds, a special feature of the cone is its association
with a colored oil droplet, which may refine color vision still further.
Visual Structure.
The
pigeon's visual system has certain features that facilitate the foraging task.
The eyes themselves are very large, filling much of the head’s interior.
While they are positioned laterally, the pupils are directed slightly forward,
providing a binocular field of view. A rich retinal structure
includes a large proportion of cones, the receptor specialized for daylight
vision. Cone pigments also provide the basis for hue discrimination. In
pigeons, as in many birds, a special feature of the cone is its association
with a colored oil droplet, which may refine color vision still further.
Another unusual characteristic of the pigeon eye
is the presence of two specialized retinal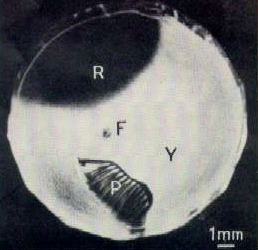 areas, each similar to the single
cone-rich human fovea. The adjacent illustration below, from Galifret (1967),
shows the retina as viewed laterally. A true foveal depression (F) is located
near, but slightly below the retina’s center. A second specialized area
(the "area dorsalis") is located above, in the upper temporal retina (R),
shown here as black. The area dorsalis lacks a depression, but is also characterized
by exceptionally dense cell structure. (The letters 'R' and 'Y' refer to
the colors that characterize the two regions; 'P' refers to the pecten,
a structure that may have one or more of several functions.) Because of
the eyes’ placement in the head, the central fovea would serve the lateral,
monocular field of view, and the area dorsalis would serve the frontal
region below the bird’s beak, including objects in the binocular field
of view.The chapter by Husband
& Shimizu (2001) considers the avian visual
system in an evolutionary context.
areas, each similar to the single
cone-rich human fovea. The adjacent illustration below, from Galifret (1967),
shows the retina as viewed laterally. A true foveal depression (F) is located
near, but slightly below the retina’s center. A second specialized area
(the "area dorsalis") is located above, in the upper temporal retina (R),
shown here as black. The area dorsalis lacks a depression, but is also characterized
by exceptionally dense cell structure. (The letters 'R' and 'Y' refer to
the colors that characterize the two regions; 'P' refers to the pecten,
a structure that may have one or more of several functions.) Because of
the eyes’ placement in the head, the central fovea would serve the lateral,
monocular field of view, and the area dorsalis would serve the frontal
region below the bird’s beak, including objects in the binocular field
of view.The chapter by Husband
& Shimizu (2001) considers the avian visual
system in an evolutionary context.
Visual function.
A number of psychophysical studies have attempted
to quantify the pigeon’s visual abilities and to compare them to humans.
Visual acuity is close to, but slightly poorer than the human’s under comparable
conditions (P. Blough, 1971; Hodos, Leibowitz, & Bonbright, 1976).
Pigeons have also proven very sensitive to hue and luminance differences.
Like humans, pigeons discriminate particularly well among lights in certain
regions of the spectrum; in particular, they discriminate best among wavelengths
we would call orange; they also show exceptional sensitivity in the "green-yellow"
region of the spectrum (summarized by Riggs, Blough, & Schafer, 1972).
An intriguing hypothesis addresses the functions
of the separate retinal areas, suggesting that the fovea and the area dorsalis
are specialized for different tasks. In observing birds in the field, for
example, it is evident that they select grain at close range as they view
the ground just below. However, when a distant observer approaches from
the side, the pigeons will show alarm without turning to face the threat.
Commenting on evidence that pigeons discriminate poorly among nearby laterally-placed
stimuli, Catania (1964) proposed that the frontal and lateral fields
are associated with different refractive characteristics; in particular,
the frontal field is near-sighted or myopic, and lateral viewing is far-sighted
or hypermetropic. This notion is attractive because it would help explain
how the pigeon can search for food and, at the same time, monitor for distant
predators. Psychophysical studies of pigeon visual acuity have provided
some support for Catania’s hypothesis; acuity for laterally-viewed targets
is better when the stimuli are relatively distant; acuity for frontally-viewed
targets improves when the targets are moved closer (Bloch, Rey, & Martinoya.
1981; P. Blough, 1971; Uhlrich, Blough & Blough, 1982).
The area dorsalis is sometimes called the "red
field." Its reddish appearance results from a relatively high concentration
of red and orange oil droplets in this region; there are proportionally
more yellow and clear droplets associated with cones in the remainder of
the retina, which appears yellow. It seems reasonable that there would
be associated differences in color sensitivity between the two retinal
areas; that is, the oil droplets may act as filters such that light reaching
the cones in the two fields would have different spectral compositions.
This hypothesis tempts an association with the pigeon’s natural tasks;
for example, Galifret (1968) noted that a red filter would enhance contrasts
between an object lying on a green background, while a yellow filter would
enhance contrasts between an object against a blue sky. A study by
Martin & Muntz (1979) provided supportive data; they revealed a depression
in sensitivity to long wavelengths when stimuli were laterally rather than
frontally presented. Other work has been less successful in documenting
effects associated with oil droplet distribution (e.g. P. Blough, 1983).
Memory Requirements
The pigeon is well prepared for the perceptual
aspects of the foraging task. However, given the varied prey and
backgrounds that it experiences, the bird also confronts strategic decisions.
Seeds differ not only in the ease with which they can be detected, but
also in their abundance, the time and effort required for their "capture,"
their size, nutritional value, and so forth. It is likely that
the needed associations are represented in a memory, which the bird would
address as it searches. Evolutionary theorists propose that the resulting
choices would be those that maximize the benefits and minimize the costs
of this behavior. A sizable literature, much of it summarized in this book,
addresses the nature of the pigeon’s memory We know, for example, the capacity
of this memory is surprisingly large (e.g. Vaughan & Greene,1983).
In their chapter in the present book, Chase and
Heinemann (2001) present a model that describes how such memories
may be constructed, stored and retrieved.
Overall, the pigeon’s perceptual and memory systems
permit it to access a large amount of information in a brief period of
time. However, foraging sometimes proceeds more efficiently when it is
possible to focus on the most relevant subset of data, even though other
aspects may be missed. Pigeons, like humans, possess the ability to redirect
their attention to restricted aspects of their perceptual or remembered
world (Blough & Blough, 1997).
The remainder of this chapter considers experimental
investigations relevant to the pigeon’s foraging behavior. The next section
considers studies of human and pigeon visual search with emphasis on common
themes is these literatures. The final section emphasizes effects on search
imposed by attentional constraints and attempts to bring together models
from naturalistic and laboratory-based approaches.
Next section: Visual Search
