Entire Set of Printable Figures For
Hierarchical Stimulus Processing by Pigeons - Cook
![]()
In one experiment, Cerella (1980) trained pigeons to discriminate intact drawings of Charlie Brown from normal drawings of other Peanuts characters. Then, Cerella reorganized Charlie Brown by altering the relations between the head, torso, and legs. He discovered that the pigeons responded to scrambled versions of Charlie Brown in the same manner as the original, intact drawings. Therefore, he concluded that the pigeon must be insensitive to global organizational properties of objects. Insensitivity to global object properties is one attribute of a particulate perceiver.
![]()
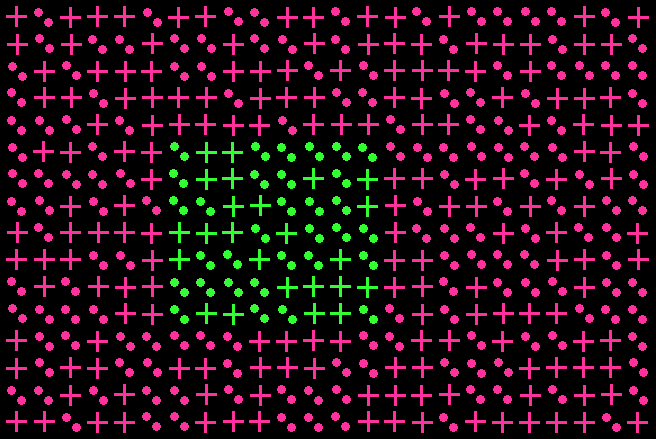
Example of Color Texture Stimulus.
![]()
Example of Shape Texture Stimulus
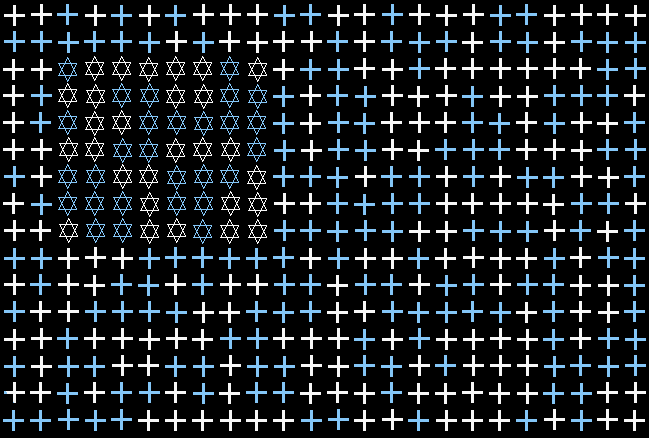
![]()
Example of Conjunctive Texture Stimulus.

![]()
Video clips can't be printed.
![]()
Example of Two-Dimensional Feature Stimulus (Color) tested by Cook et al. (1996).

![]()
Example of Two-Dimensional Feature Stimulus (Orientation) tested by Cook et al. (1996).
![]()
Example of Two-Dimensional Conjunctive Stimulus tested by Cook et al. (1996).

![]()
Example of Three-Dimensional Feature Stimulus (Color) tested by Cook et al. (1996).

![]()
Example of Three-Dimensional Feature Stimulus (Orientation) tested by Cook et al. (1996).

![]()
Example of Three-Dimensional Feature Stimulus (Size) tested by Cook et al. (1996).

![]()
Example of Three-Dimensional 1-Shared Conjunctive Stimulus tested by Cook et al. (1996).

![]()
Example of Three-Dimensional 2-Shared Conjunctive Stimulus tested by Cook et al. (1996).

![]()
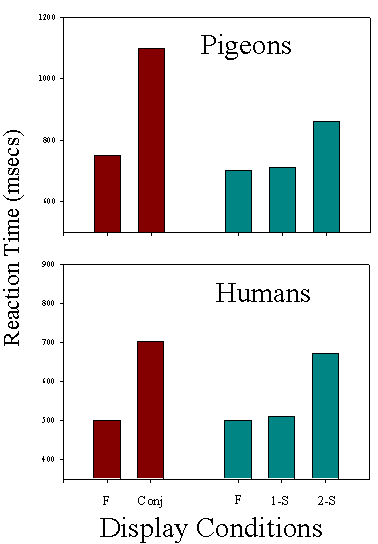
Comparison of RT measures for pigeons and humans when tested with different types of feature and conjunctive displays (Redrawn from Cook, Cavoto, & Cavoto, 1996).
![]()
Example of a Dot Density Display.

![]()
Example of a Dot Spacing Display.

![]()
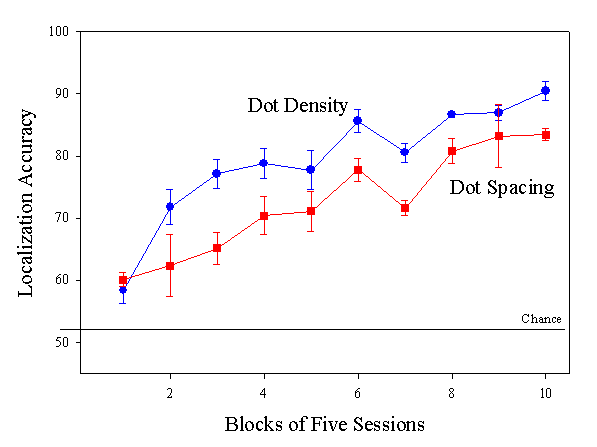
Acquisition results for four pigeons trained with dot texture displays varying in either dot density or dot.
![]()
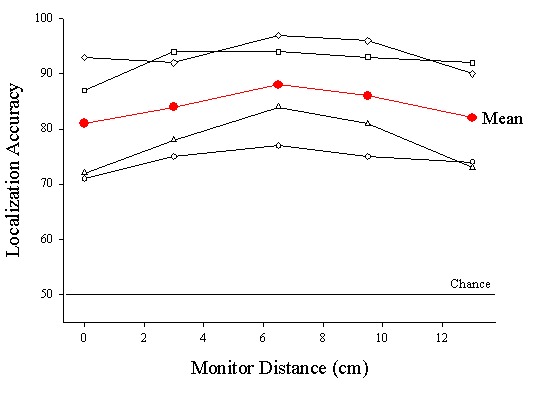
Effects of monitor distance on steady state discrimination of dot density displays. The red line is the mean performance for all four birds. The open symbols show performance for each of the four birds separately.
![]()
Animation can't be printed.
![]()
Discrimination Acquisition with Hierarchical
Stimuli and Size-Matched Controls
(Cavoto & Cook, under review).
![]()
Footnote 1.
To
interpret such a stimulus competition procedure, it is critical that the
global and local information be equally discriminable. Besides the advantages
outlined earlier, an additional benefit of training the pigeons with different
sized stimuli was that it allowed us to select stimuli in which the
discriminability (as reflected in choice accuracy) of each level was as similar
as possible. Towards this end, all probe stimuli were created from sizes
selected so that both levels supported equal levels of choice accuracy for a
particular bird. For Birds #1H and #3N this was the medium stimulus size and for
Birds #2R and #4B this was the small size, with the birds correctly
discriminating the relevant letters at the global and local levels at 84% and
86.3% respectively in sessions collected just prior to the experiment.
![]()
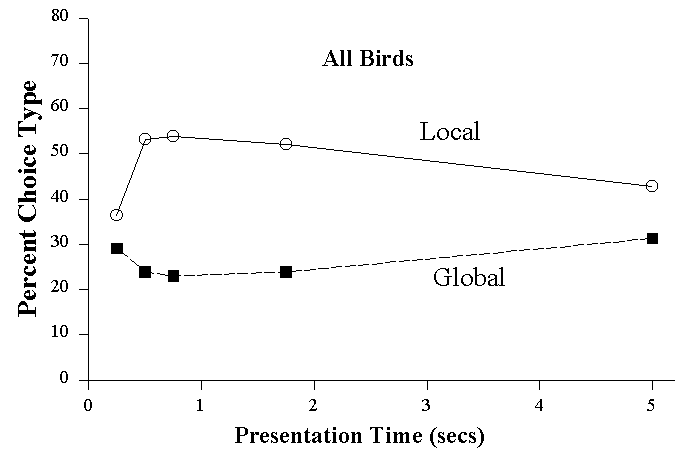
Relevant Letter Choices on Conflict Tests as a function of Presentation Time (Cavoto & Cook, 2001).
![]()
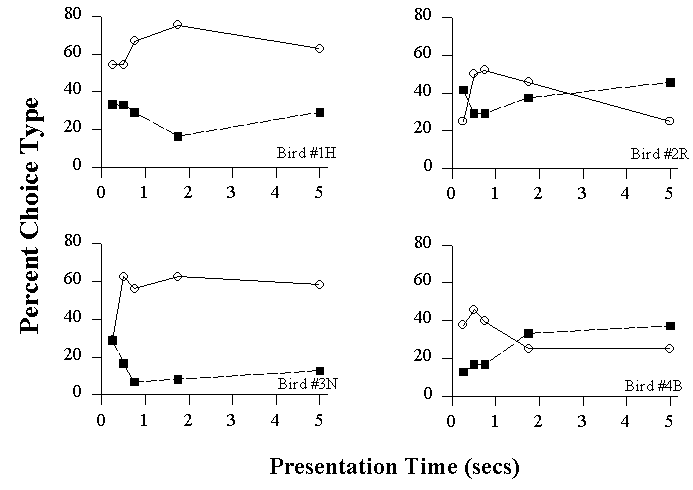
Relevant Letter Choices on Conflict Tests as a function of Presentation Time for Individual Birds.
Two birds (#1H & # 3N, the left two panels) consistently identified the local letter of the conflict stimuli more often across all stimulus durations, although this was reduced at the shortest durations. In contrast, the other two birds (#2R & #4B, the right two panels) showed systematic changes in the proportion of global and local choices as a function of stimulus duration. At the short durations, these birds reported the local letter most often, while at the longer durations the global letter was reported significantly more often. Thus, for two birds the local advantage was present across all durations, while the two other birds shifted to showing a global advantage at the longest duration (Cavoto & Cook, 2001).
![]()
Footnote 2.
In a series of experiments examining two species of nonhuman primates, Fagot and his colleagues (Deruelle & Fagot, 1998, Fagot & Deruelle, 1997; Fagot & Tomonaga, 1999) have found that baboons and chimpanzees generally exhibit an effect opposite of humans, showing a local precedence effect in which higher accuracy or faster RTs are recorded with local relevant displays, especially with sparse element arrangements. Humans tested with the same stimuli as the primates revealed the standard global precedence effect.
![]()
Figure 3. Pigeons viewed these complex stimuli with target stimuli at either the local or global level of perceptual analysis. See Figure 1 and related text in Fremouw, Herbranson, and Shimp (1998) for additional details.

![]()
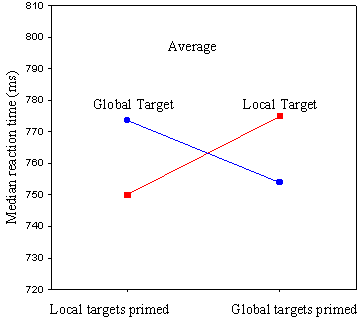
Figure 10. Pigeons’ average reaction time to recognize a target at either a local or global level of perceptual analysis, as a function of which level was primed. See Figure 2 and related text in Fremouw, Herbranson, and Shimp (1998) for additional details.
![]()
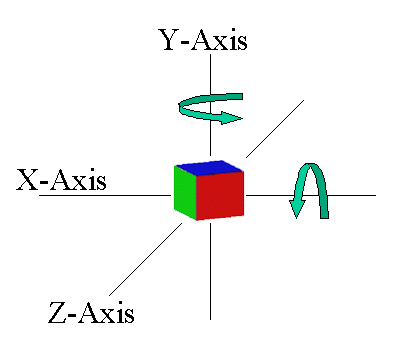
The two different axes of rotation tested in Cook & Katz (1999).
![]()
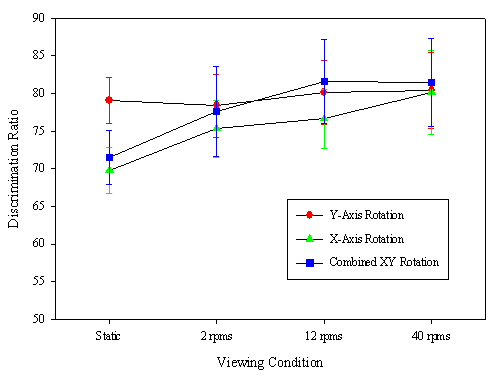
The effects of axis and rate of rotation on discrimination of cube and pyramid stimuli from Cook & Katz (1999). The red line indicates performance with Y-axis rotations of these stimuli. There was little effect of motion for this rotation, presumably because it added little new information to these unambiguous views of the stimuli. The green and blue lines show the effects of more complex motions that resulted in larger distortions in the appearance of the objects. In both of these two cases, there was a dynamic superiority effect for all three rates of rotation tested.
![]()
Examples of the "kinetic depth" stimuli tested by Cook & Katz (1999). We found evidence that three of our four pigeons were better at discriminating the structure of these "objects" when they were in motion than when they were presented at random static angles. This suggest that kinematic factors helped to recover the shape of these ambiguous images.

![]()
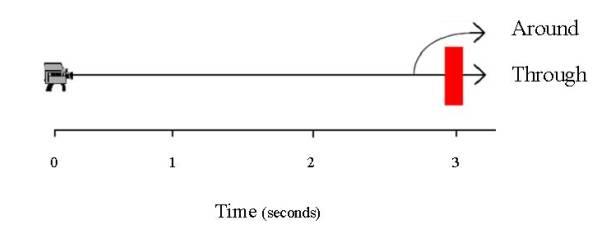
Illustration of the trajectory and timing of the two motion pathways tested by Cook, Shaw, and Blaisdell (in press).
![]()
The various objects used by Cook, Shaw and Blaisdell (in press) to used to create the two motion pathways.

![]()
Video clips can't be printed.
![]()
Mean discrimination ratios for six transfer tests with novel objects for all five successfully-trained pigeons. The red bars show transfer performance and the blue bars show baseline performance. The first four tests for the two experienced involved reintroductions of the videos not discriminated during an earlier period of training.
![]()
![]()
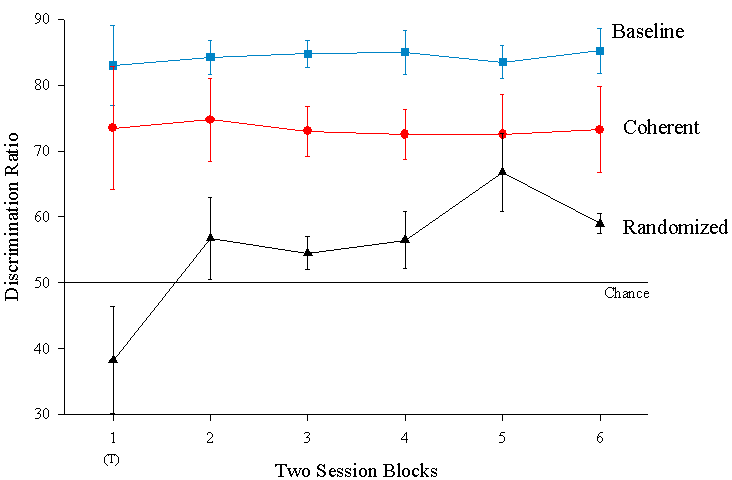
Mean discrimination ratios presented in two-session blocks for the baseline, coherent and randomized presentation conditions. The small T identifies non-reinforced transfer sessions. These results are from a our second test, in which only one fixed randomized sequence was tested, making it more comparable to the single coherent sequence tested. Only four birds were tested.