How is a Spatial
Pattern Learned and How is it Used:
Motion, Memory,
& Mapping
Clearly, spatial choices can be controlled by the spatial
relations among goal locations despite the fact that those goal locations are
not designated by any visual or other perceivable beacons, landmarks, or
geometric cues. What is the mechanism of this spatial pattern learning.
One possibility is that rats acquire response tendencies that
result in choice of poles with particular spatial relations to previously chosen
poles, and that such acquired response tendencies constitute the mechanism of
control by spatial patterns (Olthof, Sutton, Slumskie, D'Addetta, & Roberts,
1999). After being exposed to the square pattern, for example, a rat
might acquire the following response tendency: After finding two adjacent
baited poles, turn left or right and choose the next pole (Figure D-1). This rule
would result in finding a third baited pole. It requires that the rat
discriminate and remember the spatial relation between the first two baited
poles and then respond in a manner that is contingent on that relationship.
Figure D-1: An S-R explanation of correct responding
following discovery of two baited poles when pole are
baited in a square pattern
A similar (although somewhat more complex) rule could be
acquired that would increase the likelihood of choosing the fourth baited pole
in a square pattern.
The response rule that would produce control by a linear pattern
is the converse of the one that would be acquired for a square pattern: After
finding two adjacent baited poles, move in the same orientation as those two
poles and choose the next pole (Figure D-2).
Figure D-2: An S-R explanation of correct responding
following discovery of two baited poles when poles are
baited in a linear pattern
In the case of a checkerboard pattern, the description of a
response tendency explanation of control by the pattern corresponds directly to
the measure of control by the checkerboard pattern described in Part III.
That is, rats develop a tendency to make adjacent moves following choice of an
unbaited pole and/or a tendency to make skip & diagonal moves following choice
of a baited pole.
Brown, Zeiler, and John (2001) showed that the acquisition of
such response tendencies cannot explain control by spatial patterns, at least in
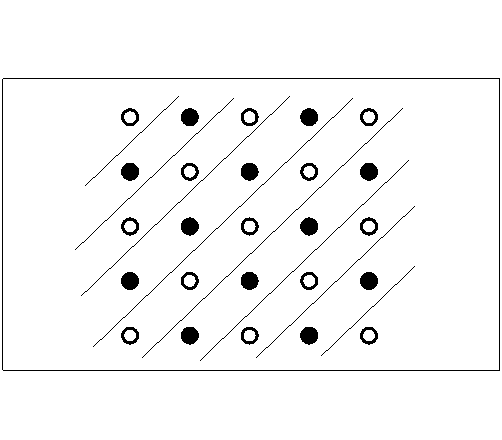
the case of the checkerboard pattern. Brown, et al. (Experiment 2) exposed
rats to a checkerboard pattern in a 5 X 5 pole box. Barriers (constructed
of plastic mesh material allowing the rats to see through the barriers)
prevented moves to poles in the same row or column (Figure D-3). Thus, without walking
around a barrier, rats could only choose poles along a diagonal axis.
|
|
Figure D-3:
One of two exemplars of the checkerboard pattern, with clear
plexiglass walls forming
diagonal alleys of poles shown. This arrangement was used
during the training phase of Brown, Zeiler, and John's (2001)
experiment. |
Thus, during this training phase, a rat could not have acquired
response tendencies that correspond to adjacent or skip moves.
Nevertheless, when the barriers were removed, the rats immediately demonstrated
a stronger tendency to make adjacent moves following choice of an unbaited pole
and a stronger tendency to make skip moves following choice of a baited pole.
This result shows that spatial relations among goal locations control behavior
in the absence of specific response tendencies.
If the mechanism of control by spatial patterns is not the
development of response tendencies, then what is it? We conclude that an
abstract representation of the spatial relations among the goal locations is
acquired. Such a representation would allow novel paths from one goal
location to another to be followed. The logic of the Brown, Zeiler, and
John's (2001) experiment and the conclusion that a flexible spatial representation
is necessary to explain the results are analogous to Tolman, Richie, and
Kalish's (1946) classic "shortcut" experiment and the resulting argument that a
flexible representation (cognitive map) of familiar locations is acquired.
However, there is a critical difference between the spatial relations involved
in the pole box task and the spatial relations among particular locations. In the case of Tolman's experiment, as in
the case of almost all more recent and current work on cognitive mapping, the locations are specific
locations in allocentric space, defined by spatial cues (e.g., beacons,
landmarks, geometric cues). In the pole box task, however, the goal
locations vary unpredictably from trial to trial. The goal locations,
therefore, must be coded in a temporary manner. The relationships
among the baited locations, on the other hand, are consistent across trials and
must be coded in a permanent manner in order to effectively control choices in
accord with the pattern. This is the sense in which the spatial pattern
must be abstract: it must be abstracted from the particular locations that are
baited on particular trials.
In order to abstract the spatial relations among goal locations,
given that the goal locations change unpredictably in allocentric space, rats
must somehow be perceiving the spatial relations among the baited poles found
during individual trials. Two possible mechanisms for doing so can be
distinguished. First, a working memory system could be used to code
the allocentric location of poles previously discovered during a trial.
The spatial relations among those locations could then be determined on the
basis of working memories for their locations. The abstracted spatial
relations among baited locations would be coded in a more permanent memory
system. According to this view, the process of spatial pattern learning is
analogous to concept learning in that the spatial relations are abstracted from
particular exemplars of baited pole locations experienced over trials.
Alternatively, a dead reckoning system could be used that
integrates the distance and direction from each baited pole discovered to the
next. According to this view, rats need not code the locations of
particular baited poles during the trial. Instead, their spatial
relationship is coded directly in terms of the vector provided by dead reckoning
as the rats moves in the pole box and chooses poles. A new vector is
initiated each time the rat discovers a baited pole. The resulting set of
vectors specifying the relations among each pair of poles forming the pattern
constitutes the learned spatial pattern.
We recently completed a series of experiments designed to
investigate the possibility that working memory for the location of previously
discovered baited poles is involved in pattern learning. To do so, we
developed pole box apparatus that allowed us to provide visual cues
corresponding to poles visited during each trial.
The video in Figure D-4 shows a trial from an experiment using the first
of two versions of this apparatus.
As the rat visits poles, a spotlight (produced from above using a data
projector) marks the location of visited poles. It was expected that such
cues would allow the rats to make fewer revisits of poles. The question
was whether it would also enhance control by the checkerboard pattern of baited
poles. If so, that would constitute evidence that working memory is also
used to keep track of where the previously discovered baited poles were, and
that these temporary memories about the elements of particular exemplars of the
pattern are involve in the ability to learned the abstracted pattern.
![]()
Figure D-4: Video of rat in in polebox with feedback stimuli indicating
chosen
arms Click image to play video
Unfortunately, the visual cues had no effect on either pole
revisits or control by the pattern. It is likely that this failure of the
cues to affect behavior was due to the brightness of the light produced by the
data projector. We suspect the the rats may not have been able to
discriminate the cues because they were masked by the ambient light produced by
the projector.
A modification of the technique used to provide visual cues
resulted in cues that did affect behavior. The base of the poles in this
second version of the apparatus (shown in Figure D-5) were constructed of translucent
material and mounted on top of holes cut in the floor of the arena. A data
projector was used to project light up from underneath the apparatus, thereby
allowing poles to be individually illuminated (as shown in Figure D-6).
|
 |
| Figure D-5: Apparatus used by Brown and
Wintersteen. Translucent base of each pole can be illuminated
from below |
|
 |
Figure D-6:
Close-up of poles in the Brown and Winterseetn apparatus. Pole
in foreground is not illuminated. Two poles in background are
illuminated. Pole illumination was used as a cue indicating
whether pole had been visited earlier during the trial. |
Brown and Wintersteen (2004) trained rats with one of the two
exemplars of the checkerboard pattern defining the location of the baited poles
on each trial. During training, the base of the pole was illuminated
whenever a rat choose the pole (or, for half of the rats, all the poles were
illuminated at the beginning of the trial and the illumination was turned off
when the rat visited a pole). During a test phase, half of the
trials did not involve use of the visual cues (the illumination of the poles did
not change). This allowed comparisons of performance with and without the
visual cues corresponding to visited locations. The visual cues did
enhance the ability of the rats to avoid revisits to poles visited earlier
during the trial. However, there was no evidence that the cues had an
effect on control by checkerboard spatial pattern. Because visual cues
corresponding to visited poles facilitated the ability of rats to avoid revisits
of those poles but had no effect on control by the checkerboard pattern, Brown
and Wintersteen argued, the working memories for pole locations used to avoid
revisits must not be involved in the acquisition of pattern learning or in the
use of learned patterns to locate baited poles. They suggested that two
separate working memory systems may be used in the pole box task: one set of
working memories reduce visits to those (previously visited) locations and a
second set of working memories code the location of previously discovered baited
poles.
Although the existence of two separate working memory systems can explain the
dissociation of memory for previous pole visits from control by spatial
patterns, another possibility is that working memory for the location of
previously visited poles is not involved in the learning or use of spatial
patterns at all. Instead, rats
could acquire the spatial pattern of baited poles using dead reckoning. Dead
reckoning could be used to detect and code the spatial relations among the poles
baited in a pattern if discovery of a baited pole defines the end of integration
of one dead reckoned vector and the beginning of another. Furthermore, the
product (vectors) of the dead reckoning process would have to be stored across
trials. Biegler (2000) has proposed an analogous function of dead
reckoning in the development of allocentric cognitive maps.
Figure D-7 shows a hypothetical path of a rat as it moves from one
baited pole to another. Note that the route of the rat might be quite
indirect and might include visits to other (non-baited) poles. The
suggestion is, however, that the discovery of a baited pole defines the end of
an instance of path integration (and the beginning of a new one). Thus,
each transition from one baited pole to another produces a vector (the arrow
should in the illustration below) that defines the spatial relationship between
those two baited poles. Each transition from one baited pole to the next
would produce one such vector.
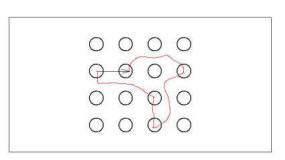 |
Figure D-7:
Illustration of path integrated spatial relation between two poles.
Rat follows path indicated in red from on pole to the other, but
codes the spatial relation between them as indicated by the arrow,
based on information provided by path integration. |
The resulting vectors would be cumulated over discoveries of baited poles both
within a trial and over trials. Over the course of trials, the square,
line, and checkerboard patterns would result in a set of vectors that code the
spatial relationships among the baited poles in the three patterns.
In Figure D-8, the vectors produced by dead reckoning are represented by
the lines connecting the (green) goal locations. The goal locations are
defined exclusively in terms of the vectors. That is, there is no coding
of the location of the goal locations in allocentric space. Their position
is learned only in relation to each other. Dead reckoning provides a
mechanism for this relational spatial coding.
Figure D-8: Hypothetical vectors (lines) representing abstracted spatial
relations
among locations (green dots). These could be formed as a result of
moving among the locations via dead reckoned spatial relations.
A weakness of this account of spatial pattern learning is that it does not
explain the ability of rats to choose in accordance with the pattern after
choosing an unbaited pole. This ability was clearly shown in the
case of a line (row) pattern by DiGello, et al. (2002). Furthermore, the
tendency to choose adjacent poles following choice of an unbaited pole in the
checkerboard pattern also indicates that rats not only learn the spatial
relationships among baited poles, but also learn the spatial relationships
between baited poles and unbaited poles. It is, of course, possible to
extrapolate the dead reckoning view of pattern learning by proposing that the
spatial relationship between all pairs of consecutively chosen poles is
discriminated used dead reckoning and that the bait status of the poles is also
coded along with the resulting set of vectors. An illustration of one
version of this idea is shown below for the checkerboard pattern. The
green nodes represent baited poles and the red nodes represent the unbaited
poles that separate the baited poles in the rows and columns of the matrix.
Learned vectors define the relations among the baited locations and between the
adjacent baited and unbaited locations.
Figure D-9: Hypothetical vectors (lines) representing abstracted spatial
relations
among two kinds of locations - baited (green dots) and unbaited (red dots).
Thus, dead reckoning provides a potential mechanism for abstracting the spatial
relations that control choices in the pole box. We do not yet have any
direct evidence for or against the possibility that dead reckoning is providing
this information. Thus, abstraction of the spatial relations among baited
poles may be mediated either by working memories for the particular locations
baited on individual trials or by dead reckoning.