|
Visual Control of Sexual Behavior Chana K. Akins
& Melissa Burns |
![]()
|
|
I. Introduction Reproductive behavior is central to the ultimate fitness of organisms. Various sensory systems play a role in regulating reproductive behavior. The predominant system is typically highly-developed and species specific. For example, the olfactory system is highly developed in rats and serves as the primary sensory system involved in the reproductive behavior of rats. In contrast, the auditory system is highly developed in songbirds and it plays a critical role in social and courtship interactions. Just as the auditory system is highly developed in songbirds, the visual system is highly developed in some gallinaceous species of bird and serves as the primary sensory system involved in the reproductive behavior of birds. The purpose of this chapter is to review the literature on the various types of visual cues that affect sexual behavior and sexual learning and on how these cues influence reproductive behavior. The first section of this chapter is a description of the predictive and synchronizing cues used by avian species in naturalistic settings that are associated with the visual system and that are critical for the performance of copulatory behaviors. Predictive cues are preparatory cues that serve to set up reproductive condition. They primarily serve to induce reproductive readiness and to activate the physiological mechanisms that regulate sexual behavior. Predictive cues include photostimulation, the hypothalamus-pituitary-gonadal axis, and sex hormones. In contrast, synchronizing cues serve to elicit reproductive behavior and may include sexually dimorphic plumage, ornaments, and courtship rituals. These cues serve to enhance sexual responding in females of many bird species. The second part of this chapter is a review of the laboratory research that has been conducted to examine various types of visual cues. It includes a description of how these visual cues facilitate both unlearned and learned sexual behavior. Visual cues that have been found to enhance unlearned sexual behavior include species-specific female cues, while those that have been shown to facilitate learned sexual behavior include species-specific female cues, local or discrete cues, and environmental contextual cues. Because we want to present an integrated account of both unlearned and
learned sexual behavior, much of our discussion in the latter half of this
chapter will include research that has been conducted with domesticated
Japanese quail (Coturnix japonica). Domesticated quail are
particularly well-suited for studying sexual behavior, in part, because II. Visual Cues for Successful Mating The survival of a species is dependent on the members of that species reproducing. For such an important function, however, reproduction is quite complex. To breed successfully, birds need to predict the onset of the breeding season, travel to a suitable breeding site, develop and maintain functional gonads, establish and defend a territory at the correct time, select and/or seduce a mate, coordinate sexual behavior with the mate, and switch from sexual to parental behavior when it is appropriate to do so. A large repertoire of diverse behaviors is required to navigate through each step of the reproductive process. The wrong behavior at the wrong time could prove to be detrimental to the reproductive effort. Fortunately, there are many cues that help to direct the reproductive effort along the correct path. Some cues come in the form of long-term changes throughout the environment, such as an increase in temperature and day length as spring draws near. Such omnipresent cues tend to cause relatively long-lasting internal changes in birds, such as an elevation of certain sex hormones that last throughout the breeding season. Ambient cues help birds to predict when the breeding season will begin and when it will end. Other external signs occur very quickly and in a limited area in the environment, such the appearance of a reproductively mature conspecific on one�s territory or the sound of a neighboring male singing to defend his territory. These types of discrete cues can elicit sudden changes in the bird�s internal state, such as a brief surge in the release of sex hormones. Discrete cues often aid in the synchronization of mating behavior. Thus, it is important, even vital, that each reproducing member of the species be highly tuned to its external and internal environment in order to detect cues that signal when, where, and towards whom each specific behavior should occur. Researchers have identified many environmental signals that serve as predictive or synchronizing cues for avian species. Many of these cues are of an auditory nature (e.g., vocalizations), some are somasthetic (e.g., humidity and temperature), but most of the cues used by avian species are visual. The present chapter, therefore, focuses on cues that are of a visual nature. It is not a simple matter to turn a visual signal into reproductive activity. Rather, the conversion of external cues into sexual behavior is carried out by a series of neural and physiological events. In order for an environmental cue, such as the presence of a potential mate or the change in day length, to affect the reproductive physiology of a bird, the cue must be perceived by the nervous system, converted to meaningful information, and directed towards a target area in the body. Photic energy from the environment enters the avian brain both through the eyes and by penetrating the skull to activate extra-retinal photoreceptors in the brain. Discrete visual cues in the environment, such as a brightly colored patch of feathers on a conspecific, enter the nervous system through the retina of the eyes. Ambient visual cues, such as daylight, activate photosensitive loci in the brain both indirectly, through the eyes, and directly, through the skull. The hypothalamus of the bird brain contains special cells that are sensitive to extremely low light levels, intensities comparable to the amount of light that can penetrate brain tissue. The hypothalamus also receives one-way afferent connections directly from the retina. These fibers terminate primarily in the suprachiasmatic nucleus, an area of the anterior hypothalamus that is essential in gonadotropic ("gonad changing") mechanisms in birds (doves - Cooper, Pickard, & Silver, 1983; ducks � Bons, 1976; jackdaw � Meier, 1973; Java sparrow � Ebihara & Kawamura, 1981; pigeon � Meier, 1973; Reperant, 1973; and house sparrow � Hartwig, 1974). Many neurons in the avian hypothalamus are specialized for the synthesis and secretion of peptides into the bloodstream. Once in circulation, these peptides act as hormones on distant cells. Their effects are enduring and serve to control neuron excitability and synaptic effectiveness. Thus, peptides can enhance or suppress neuronal sensitivity to specific environmental stimuli. These long-lasting activities are thought to be important for a variety of behavioral functions, including mood, motivational state, and learning. The hypothalamus also controls the endocrine system. Regulatory hormones produced by the hypothalamus are secreted into a blood vessel, the hypophyseal portal blood vessel, which drains into the anterior pituitary gland. The pituitary gland produces specialized hormones that act on target glands and tissues in other parts of the body by prompting the release or inhibition of certain hormones. Within the reproductive system, stimulation of the hypothalamus results in the secretion of gonadotropin-releasing hormone (GnRH) into the hypophyseal portal blood vessel. When activated by GnRH, the anterior pituitary secretes two gonadotropin hormones, follicle-stimulating hormone (FSH) and luteinizing hormone (LH). The gonads detect the presence of gonadotropins in the blood stream and respond by producing gametes and sex hormones. The sex hormones then bind to hormone receptors sites in specific loci in the brain (e.g., the pre-optic area, nucleus preopticus paraventricularis manocellularis, and nucleus hypothalami lateralis). Stimulation of these brain regions generates synaptic transmissions that ultimately create either changes in reproductive physiology, such as the development of song regions in the brain, or a specific motor activity, such as erecting and spreading one�s beautifully ornate tail feathers in the presence of a female. The environmental cues most important for successful breeding are those that are most highly correlated with the breeding season. Because breeding activity occurs at approximately the same time each year, the diurnal photocycle, or varying light:dark ratio during the year, is an excellent indicator of when to prepare for the breeding season and when to terminate breeding. Testicular growth and hormone production are strongly correlated with day-length ("photoperiod"). Long days of the summer breeding season support large, productive testes while short days of winter instigate atrophy and general inactivity of the testes (Wingfield & Farner, 1993). In this respect, changes in the photoperiod provide predictive information (Wingfield, 1980) that induces gonadal maturation prior to the breeding season and causes degeneration of gonads at the end of the breeding season. Since the effects of day length on gonadal development were first reported by Rowan in 1925, the phenomenon has been demonstrated in about 70 species. The development of a functional reproductive system requires several
weeks, thus individuals must predict the ensuing reproductive period and
begin maturation well in advance so that the gonads are functional when
environmental conditions become favorable for breeding. This process has
been well investigated in the males of many species (e.g.,
Mattocks, Farner, & Follet, 1976; Donham, 1979; Assenmacher
& Jallegeas, 1980; Bluhm, Phillips, Burke, & Gupta, 1983;). As day
length increases, photic stimulation of the hypothalamus results in the
secretion of GnRH. When activated by GnRH, the anterior pituitary secretes
two gonadotropin hormones, follicle-stimulating hormone In general, wild avian species maintained on natural day lengths show a gradual increase in testes volume and plasma levels of sex hormones in the spring. These hormone levels remain high until midsummer and then decline. The testes also show a marked decrease in size as day-length decreases. In most species, the significant elevations of circulating sex hormones in the spring coincide not only with development of the testes and secondary sexual characteristics, but also with heightened aggression as territories are formed, and with periods of heightened sexual behavior (Mattocks, et al., 1976; Donham, 1979; Assenmacher & Jallegeas, 1980; Bluhm, et al., 1983; Stokkan & Sharp, 1980; Sharp & Moss, 1981; Scanes, 1974; Wilson & Follet, 1974).The importance of the eyes relative to the extra-retinal photoreceptors in controlling photo-gonadal mechanisms is poorly understood. Studies have shown that when the eyes are removed, some bird species still show photoperiodically induced gonadal growth (Benoit, 1938; Beniot & Assenmacher, 1966; Menaker, Roberts, Elliot, & Underwood, 1970). For instance, White-crowned sparrows have light receptors in the ventromedial hypothalamus. Pinpoint illumination of these hypothalamic receptors by means of a single, thin light-conducting optical fiber induces both testicular growth and migratory behavior (Yokoyama & Farner, 1978). However, other species, such as domesticated quail fail to show gonadal responses to changes in photoperiod when blinded by bilateral enucleation (Bons, Jallageas, & Assenmacher, 1975). Thus it seems that the relative contribution of the retina versus extra-retinal photoreceptors to photoregulation in intact birds is diverse. Other visual cues that are critical for successful breeding are those provided by potential mates or nest-related stimuli. These cues tend to be discrete rather than ambient cues in the environment. Although the details of the neural processing of discrete visual cues remain unclear, it is likely that the cues enter the nervous system through the retina of the eyes and follow the retinohypothalamic pathway to the hypothalamus. The hypothalamus then acts to indirectly increase or decrease sex hormone secretion. These hormones act on specific locations in the brain which, in turn, activate particular motor responses related to reproduction. For instance, many cues provided by potential mates are highly effective in stimulating LH release and they produce quick-acting changes in sexual behavior. Unlike photic stimulation which causes slow and long-lasting changes to reproductive physiology, synchronizing cues may aid a bird in focusing its reproductive efforts on a potential mate, thereby releasing sexual behavior at the appropriate time during the breeding season. Discrete cues such as the presence of a nest box, nesting material, or a potential mate, are more closely related in space and time to reproductive activity than the light cycle. Due to their close spatial and temporal relationship, discrete cues can be powerful stimuli that engender specific reproductive behaviors to help synchronize the reproductive effort of the pair. In most species, initial predictive information, such as photoperiod, is necessary to bring a bird into reproductive condition in time for the breeding season. Specific stimuli then predominate to engage the readied bird in sexual behavior with an appropriate mate. Synchronizing cues may be in the form of nesting material, as is the case of the cockatiel (Shields, Yamatoto, & Millam, 1989), or they may be in the form of species-typical and/or sex specific characteristics of a potential mate. For example, plumage and courtship dances are very common in birds. In fact, male birds are notorious for signaling sexual interest with visual displays. Other visual signals involve more subtle behaviors that occur in the course of reproductive activities. Many synchronizing signals are not obvious to human observers. For instance, it was long believed that female birds choose mates based on male characteristics that are visible to the human eye. Recent research has shown that this is not always the case. Rather, some species of birds make mate choices based on ultraviolet plumage coloration to which humans are blind (Bennett, Cuthill, Partridge, & Maier, 1996; Bennett, Cuthill, Partridge, Lunau, 1997). However, many sexual displays involve robust behaviors and brightly colored plumage and ornamentation that are easily discernible by the human eye. These the sexual displays have been well studied and provide wonderful examples of how birds can use visual cues to sexually stimulate members of the opposite sex. Plumage is the most obvious indicator of species, sex, reproductive maturity, reproductive condition, dominance, and health. However, male birds also display elaborate ornamentation, engage in courtship behavior and build structures to attract females. The most effective form of visual stimuli in eliciting sexual behavior varies among species of birds. Elaborate ornamentation of males of some species serves as a strong sexual stimulus for females. Types of ornamentation include colorful or iridescent body feathers, crests, elongated or elaborate tails, and variable large areas of colorful skin exposed on the head, especially around the eyes. Often, the more exaggerated the trait, the more likely it will elicit copulation from the opposite sex. For instance, long-tailed widowbirds can have tails up to 0.5 meters long. This is a very attractive attribute since female long-tailed widowbirds typically prefer males that have the longest tails. When males� tails were artificially enhanced or shortened by an experimenter, females were observed to prefer males with "super tails" that were 25 cm longer than natural (Andersson, 1982). Playing on the relationship between exaggerated secondary sex characteristics and sexual success, males often posture to emphasize ornamental feathers and color patches.
Peafowl offer an excellent example of a feature display. Colorful iridescent eyelike ocelli are located on the highly elongated tail coverts; about 100-175 such ocelli may be present on a single adult male. These male displays are mostly frontally oriented to exhibit the ocelli maximally. In an observational study, Rands, Ridley, and Lelliot (1984) found that male peafowl that were reproductively successful were of intermediate age, did not have longer tails or greater body mass than other males, but they spent more time displaying sexually. Furthermore, the males with the most elaborate trains were typically most effective in attracting females (Petrie, Halliday, & Sander, 1991). Thus, the beautifully iridescent ocelli of the peacock tail acts as a sexual stimulus for peahen. The Bulwer�s pheasant also signals females of sexual interest by displaying a fan of tail feathers. The stark white tail feathers contrast dramatically with the dark black feathers on the body. Much like peafowl, the male great argus pheasant signals his sexual interest in a female by fanning large, iridescent feathers. However, the ornate feathers are located on his wings instead of his tail. Male great egrets also attract sexual partners by displaying special plumage. Their sexual signal involves the erection of long white filigree feathers located on the back and tail. The male lyrebird has a greatly elaborated version of the great egret display. Lyrebirds have similar long white filigree feathers in the tail. When displaying, males emphasize their special plumage by throwing the tail feathers forward over the back and head.
For female great frigate birds, the most powerful sexual stimulus comes in the form of an inflated red throat. Male frigate birds solicit females from a nest site by inflating a fleshy scarlet pouch beneath the throat. When a female is nearby, males accentuate their ballooning throats by spreading and vibrating their black wings. The kori bustard also inflates a throat pouch to attract females, however, the ballooning pouch is covered in long, wispy feathers rather than the brightly colored flesh of the frigate bird. For several species of New Guinea birds, sexual stimuli take the form of plumage displays that leave males looking less like birds and more like abstract works of art. (See the Black sicklebill, Wilson�s bird of paradise, Parotia bird of paradise, and 12-Wired bird of paradise). For each species of bird, only a specific shape, angle, color, or movement that is associated with the males� display sexually stimulates the female. The male of the species that is best able to match the females� prototype stimulus will be the most successful at mating.
Courtship dances are another type of visual signal used to induce sexual behavior in potential mates. For instance, the sexual display of the great snipe male involves vigorous activity rather than ornamentation. When a female approaches a male, a male will shift his activity from territorial defensive behaviors to intense flutter-leaping near the female. Females of lekking species are sexually stimulated by the sight of a gathering of males vigorously displaying their most sexually stimulating attribute. The conservatively dressed male buff-breasted sandpipers gather in northern Canada to breed. When a female arrives at the lekking ground, the males will display their best attribute � a snow white armpit � by raising one wing. When under close scrutiny by a female, the male may become excited and expose both armpits. This display is very appealing to female buff-breasted sandpipers. In the case of the Lesser Prairie Chicken, ornamentation and activity are combined to stimulate potential mates. About 14 sexually mature males gather on a common lekking ground to display. The males produce a yodeling vocalization, which exposes and expands brightly colored skin patches at each side of the neck, all the while fanning and closing the tail feathers. When a female approaches the lek, a new behavior is performed at high intensity, the "nuptial-bow" posture, which is often followed by copulation (Johnsgard, 1994). Count Raggi�s bird of paradise is another lekking species with a spectacular sexual display. Males attract potential mates by bending forward and vigorously flapping their wings while fluffing out long colorful plumage on their backs . The male Raggiana bird of paradise also uses high-intensity activity to accentuate his beautiful plumage.Temmick�s Tragopans have a spectacular courtship display. Males of this colorful species drum their wings rhythmically while hiding behind a rock or fallen log. When a female approaches, the male springs to the top of the rock or log then erects a blue feather horn above each eye, fluffs his body feathers, and spreads his wing feathers in a downward position. He will hold the pose for several seconds, then run to the location from which the female observed his display. If she is still there, they will copulate. It is hypothesized that the display serves to mesmerize the female into temporary immobility (Johnsgard, 1994). Regardless of the effect on the female, it is evident that surprise is a vital part of the display. When captive males are not provided with a suitable location in which to hide and display, courtship may not take place at all (Johnsgard, 1994). Building complex, highly decorated, colorful structures allows the
visual display to exist independently of the male, freeing him from
burdensome ornamentation and bright colors that increase risk of
predation. This is the method of display employed by bower birds.
Males of this In some species of birds, the elaborate and colorful male displays will prompt a female into nest building activity or receptive squatting. In other species, the particularly enthusiastic hopping and bobbing by the male is sufficient to bond a female with that male for the remainder of the breeding season. Regardless of the form it takes, the result of an effective sexual displays is cooperative copulation between the pair. In summary, the first section of the chapter has provided descriptions and examples of various visual stimuli and their role in eliciting and regulating courtship and sexual behavior in a naturalistic setting. Although naturalistic observations provide valuable and interesting information about courtship and sexual behavior, they do not provide information about the causes of these responses with regard to visual stimuli. Laboratory experiments allow for experimental manipulation and control of various stimuli to better ascertain what visual stimuli may be involved in the regulation of sexual responding. Such manipulations invariably involve introducing circumstances that are not likely to occur in the field. This research is both necessary and valuable because it can serve to identify sources of visual stimuli that control sexual behavior and the mechanisms that accompany them. The next section of this chapter provides a review of some laboratory experiments that have been conducted to investigate visual control of unconditioned sexual behavior. The techniques used to investigate visual control of sexual behavior involve presenting taxidermic models that contain various visual features of a female quail. The effectiveness of visual features in eliciting sexual responding may then be assessed by monitoring sexual responding of male quail. Some of the experiments involve measuring a social proximity response while others involve monitoring more direct indices of sexual responding such as the frequency of grabs, mounts and cloacal contact movements. III. Visual Control of Sexual Behavior in the Laboratory Laboratory Techniques for Isolating Visual Stimuli One laboratory technique that has been used to identify socially
effective visual stimuli involves using a window test The social proximity behavior of male quail is thought to share characteristics with mate guarding in other birds. Mate guarding is primarily controlled by the visual cues of a female conspecific (Lumpkin, Kessel, Zenone, & Erickson, 1982). Males of some avian species, such as magpies (Birkhead, 1979) and bank swallows (Beecher & Beecher, 1979) display social proximity by remaining near a female conspecific. The social proximity behavior of male domesticated quail is similar to the mate guarding behavior of other avian species, except that, it does not appear to be strongly influenced by the reproductive state of the female conspecific (Domjan & Hall, 1986). It does, however, appear to be controlled primarily by the visual cues of the female (Domjan & Hall, 1986; Domjan & Nash, 1988). Another technique that has been useful in identifying the critical visual stimuli that control social and sexual behavior is the use of taxidermically-prepared models. Live stimulus animals present a complex of visual, auditory, olfactory, and behavioral stimuli, any of which could potentially elicit a sexual response by a conspecific. Use of taxidermic models is an effective technique for isolating visual stimulus features. Taxidermic models of females are effective in eliciting male copulatory behavior in various galliform birds, including quail (Domjan, Greene, & North, 1989), chickens (Fisher & Hale, 1957; Carbaugh, Schein, & Hale, 1962), and turkeys (Schein & Hale, 1957, 1965). Using the window test apparatus and taxidermically-prepared models of male and female quail, Domjan and Nash (1988) evaluated whether the static visual features of a female model were sufficient to control male social proximity behavior in the absence of vocalization, olfactory cues and movement. They found that male quail that had visual access to a normal adult female spent more time near the opened window than when they had visual access to an empty side cage or one containing live male birds. More importantly, the social proximity behavior of the males presented with female models was similar to the social proximity behavior to a live female quail. Thus, the female�s calls and movements were not necessary to stimulate social proximity behavior in males. Species-specific Female Cues: Single-stimulus Tests of Social Proximity Partial taxidermic models are useful in testing the effectiveness of various female conspecific body parts in eliciting male copulatory behavior. Domjan and Nash (1988) investigated the social proximity behavior of male quail to taxidermically-prepared models of male and female quail with various body parts. Males were given single-stimulus tests with the following models in this order: a full-body female quail model, a full-body male quail model, a female quail head and neck, a male quail head and neck, a full-body female quail model whose head was covered with a brown cloth hood, a female quail head and neck suspended upside down, a full-body male quail model, and a full-body female quail model. The amount of time subjects spent near a window through which they could see a model was recorded. (See previous section on Common Techniques for Isolating Visual Stimuli for more detail.)
Species-specific Female Cues: Single-stimulus vs. Choice Tests of Copulatory Behavior Male turkeys respond best to models that contain a female�s head and neck (Schein & Hale, 1957, 1965; Schoettle & Schein, 1959), but male chickens respond best to models that contain a female�s body (Carbaugh et al., 1962). Schein and Hale (1965) speculated that the differential effectiveness of cues provided by the body of a female for male turkeys and chickens is related to the postures involved in their copulatory behavior. After mounting, a male turkey sees only the female�s head and neck because he completely covers the rest of her body. In contrast, a male chicken maintains visual contact with the female�s body during copulation. A male chicken grabs the hen�s neck or comb at the start of the copulatory sequence and maintains the grab during the mount and cloacal contact movements, ensuring visual orientation to the hen�s body. The copulatory behavior of male domesticated quail is similar to that
of chickens. A male quail first grabs the feathers of the
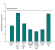
The choice tests supported different conclusions about the relative
contribution of various body parts in controlling male copulatory behavior
(See Figure 4). More
copulatory responses were directed toward the whole model than toward the
head model. In addition, more copulatory responses were directed toward It�s unclear why the choice tests were more effective in revealing control of copulatory behavior by the female�s body, rather than by her head and neck features. One possibility is that choice tests are more effective in revealing control by weak stimuli. Perhaps the cues provided by a female quail�s body are relatively weak compared to her head and neck cues, but when two models are presented, body cues still influence sexual responding. In summary, the findings of the laboratory experiments that have been conducted to investigate the stimulus control of visual features on male sexual responding indicate that the species-specific cues of the female, specifically the head and neck features, elicit more social proximity behavior and more copulatory responding than other visual features. (The exception to this appears to be when animals are given a choice between copulating with a model that contains head and neck cues versus a model that contains body cues.) That species-specific cues facilitate sexual responding may not be coincidental. As previously discussed (see introduction), male and female quail are sexually dimorphic and the head and neck features are the most sexually dimorphic plumage traits of Japanese quail. These visual stimuli appear to be the most important visual stimuli in eliciting sexual responding. The findings of these laboratory studies are in accordance with naturalistic observations in which species-specific cues and sex-specific stimuli are important in synchronizing a mating event. Given that sexual behavior is subject to the influence of prior experience, it is necessary to investigate aspects of sexual behavior that result specifically from prior experience with visual stimuli. The experiments in the previous section did not involve manipulating experience of the subjects to visual stimuli. Therefore, the next section will review several laboratory experiments in which the acquisition of sexual responding to various visual stimuli is investigated. Species-specific female cues, local cues, and contextual cues will be used as visual stimuli. IV. Visual Control of Learned Sexual Behavior in the Laboratory Pavlovian Conditioning of Visual Stimuli
In an attempt to identify the species-specific cues of females, investigators have conducted experiments with sexually experienced subjects. Less experienced subjects do not respond as reliably as sexually experienced ones (e.g., Fisher & Hale, 1957). Sexual experience can enhance various aspects of reproductive behavior. For example, sexually experienced ring doves (Lehrman & Wortis, 1960) and domestic chickens (Fisher & Hale, 1957) are more successful in their reproductive behavior than are sexually inexperienced birds. These results suggest that subjects may learn something over the course of sexual experience. Learning may occur in the form of Pavlovian conditioning. In some of the first Pavlovian conditioning experiments, Wolfsohn in 1897 in Pavlov�s laboratory, investigated the salivary response of dogs to the visual cues of materials such as sand (see Domjan, 1993). Initially, the visual cues of the sand were ineffective in eliciting salivation; only the tactile cues of sand in the mouth worked. However, with repeated experience with sand in the mouth, the previously ineffective visual cues of the sand (conditioned stimuli) became associated with the biologically significant tactile cues of the sand (unconditioned stimuli). Similarly, in sexual behavior, only some of the visual features of a potential sexual partner or mate may be effective in eliciting sexual behavior initially. With repeated sexual experience, initially ineffective features of a sexual partner may become associated with the biologically significant event, copulation, and thereby come to elicit sexually relevant responses. This suggests that animals may learn about the visual features of potential sexual partners and that what they learn may determine the effectiveness of visual cues in eliciting future social behavior. Just as in naturalistic observations, visual stimuli that are
particularly effective in eliciting learned sexual behavior often involve
stimuli that contain species-specific cues. Studies with domesticated
quail have indicated that the sexual response of male quail to female
conspecifics is, at least, in part due to what males learn about the
species-specific cues of the female quail. Cusato and Domjan (1998)
recently elucidated on the significance of species-specific female cues in
sexual learning. In their experiment, two CSs were used; a taxidermically-prepared
female head and partial Evidence from several studies indicates that arbitrary local visual cues may also come to control sexual responding. In a previous experiment (Hollis, Cadieux, & Colbert, 1989) with male blue gourami fish (Trichogaster trichopterus), male and female gouramis were presented with a 10 sec red light CS followed by a 15 sec exposure to each other (the US). Both male and females immediately approached the CS and engaged in a frontal display involving the erection of all fins. In addition, classically conditioned males significantly attenuated their territorial aggression and began courtship activities almost immediately. Thus, reproductive benefits might result from the Pavlovian conditioning of local visual cues with copulatory opportunity. In a similar experiment (Domjan, Lyons, North, & Bruell, 1986) with
birds, male quail were presented with a 10 sec red light, the CS, followed
by the opening of an opaque door that separated the male�s test cage
with that of a female�s cage. Male quail were allowed five min to
copulate with the female bird in the side cage. A control group was given
copulatory access to a female bird 3-5 hours after the red light was
presented. The amount of time subjects spent near the red light was
measured on successive trials for 25 trials. Evidence for Pavlovian sexual
conditioning was Similar results have been reported with a more complex visual stimulus,
a stuffed toy dog (Domjan, O�Vary, & Greene, 1988). The toy measured
about 14 cm from the tail to the tip of its nose. It had a bright yellow
terry cloth surface, a thin black collar, black nose button, and white
eyes with small black pupils. Male quail in the experimental group were
presented with the toy for 30 sec followed by copulatory opportunity with
a female quail. Quail in the control group were presented with the toy for
30 sec, 90 min after copulatory opportunity with a female quail. Two The most complex visual stimuli involved in conditioning are contextual cues or spatial cues. Not only do they include a wide variety of visual stimuli, but they may also contain local and/or species-specific visual cues. When contextual cues contain local and species-specific visual cues, they serve to modulate sexual responding. Context modulation of local visual cues has been examined in studies with domesticated quail. In one study (Domjan, Akins, & Vandergriff, 1992), male quail were exposed to two distinct contexts that both contained a square holding box (the local CS). One context had a sand floor, white side and back walls, a wire mesh front wall, and contained dried tree branches. The other context contained a wire-mesh floor, natural wood-grain side walls, clear plastic front wall, and contained no tree branches. Subjects were housed in each context on alternate days, so that they spent an equal amount of time in both places. Exposure to the two contexts differed only in that a female bird was released from the holding box in one context (the sexual context) but not in the other context (the nonsexual context). Male quail in the sexual context, therefore, were allowed to copulate with the female quail that were released from the holding box in the sexual context. Results indicated that subjects increased the amount of time they spent
near the holding box across trials in the sexual In addition to modulating approach responses to local visual stimuli, context modulates approach behavior to the species-specific cues of female quail. In the same experiment described above (Domjan et al., 1992), after male quail were given 20 trials in the sexual context and 20 trials in the nonsexual context, they were tested for their response to a female quail in each context. Figure 9 shows that the presence of the female bird stimulated significantly more approach behavior to theContextual cues may also form a direct association with a sexual US. The difficulty in testing for direct context-US relationships in sexual learning is that the sexually conditioned response is usually directed toward an object in the context. The object, thereby, becomes the CS and context presumably modulates responding toward it. A recent experiment (Akins, 1998) was conducted to investigate the
direct association of context with a sexual US by using a place preference
procedure. An initial preference test was conducted to determine which of
two distinct contexts
male quail preferred. One context had green walls and a
wiremesh floor while the other context had orange walls and a sand floor.
Subsequently, the quail were given several trials in which they received
copulatory opportunity with a female in their nonprefer V. Theoretical Implications: The Behavior Systems Approach
The control of the sexually conditioned response by visual cues fits nicely with a behavior systems approach (see Timberlake, 1994 for review). According to this perspective, behavior systems consist of coordinated sets of responses that are required to accomplish a biologically important task. In a species with a well-developed visual system, such as the domesticated quail, the response the bird makes may depend on the presence or absence of certain visual cues, as well as their temporal and spatial relationships. For example, the absence of a female quail may elicit increased locomotor activity or general search behavior for a potential sexual partner. When local visual cues signal that a potential partner is nearby, approach behavior or focal search behavior is elicited. When the male sees the head and neck of the female, he may approach and then copulate with the female. Thus, the behavior systems approach can be used to predict the nature of the conditioned response with respect to the type of visual stimuli used to elicit the response.
Successful reproductive behavior requires the presence of both predictive and synchronizing cues. Environmental cues such as daylength or photoperiod serve as predictive cues that are often correlated with the physiological mechanisms, such as gonadal maturation and hormone production, that further regulate reproductive behavior. Although, the role of the visual system in controlling photo-gonadal mechanisms is not completely understood, there is some evidence for the influence of visual stimuli on the hypothalamus. In contrast to predictive environmental and physiological cues, other visual stimuli may serve as synchronizing cues that influence reproductive behavior more immediately. Synchronizing cues that release sexual behavior include decorative ornaments, elaborate courtship rituals, the building of intricate structures, or a combination of these cues. Naturalistic observations have provided a plethora of examples that implicate the importance of the visual system in regulating courtship but they do not provide causal information. Laboratory studies allow for the manipulation and control of stimulus presentation. The findings of laboratory studies conducted to investigate which visual features of the female are important for eliciting sexual behavior indicate that the species-specific cues of the female are most important. In male domesticated quail, head and neck female features stimulate as much social proximity behavior as the full body of a female bird. The head and neck cues of the birds may be effective visual stimuli in eliciting male sexual responding because of their high degree of sexual dimorphism. In contrast, when given a choice, males direct as much copulatory responding to a model that contains no head and neck cues as they do to a model with head and neck cues. Thus, there may be circumstances in which male quail use other visual features of the female during sexual responding. Learned sexual responses are similar in form to unlearned sexual responses except that the nature of learned sexual response may be even more tightly controlled by the type of visual stimuli that elicit them (see Domjan, 1994 for review). Typically, female species-specific cues come to elicit conditioned approach and conditioned copulatory responses whereas arbitrary local cues come to elicit conditioned approach but rarely conditioned copulation. Sexually conditioned contextual cues serve to modulate species-specific and local cues. They enhance conditioned copulatory responding toward species-specific cues and they modulate conditioned approach behavior toward local cues by providing discriminatory cues that signal when copulatory opportunity will become available. From a functional perspective, conditioning to visual stimuli has been shown to contribute to the reproductive fitness of organisms. For example, not only does signaling the accessibility of a female blue gourami fish enable classically conditioned male blue gourami�s to attenuate their initial aggressive response toward arriving females, but more importantly, classically conditioned males were able to spawn with females sooner, clasp females more often, and produce more young than males that did not have the benefit of a signal (Hollis, Pharr, Dumas, Britton, & Field, 1997). Similarly, Domjan, Blebois, & Williams (1998) found that after several pairings of a distinct environment with copulatory opportunity, a stimulus that contained some female head and neck features presented in the environment resulted in the release of greater semen volumes and greater numbers of spermatozoa in male quail. The findings of these experiments and others suggest that understanding the role of function is critical to our understanding of animal learning and cognition (see Hollis, 1997 for review). Akins, C. K. (1998). Contextual excitation and modulation of conditioned sexual behavior. Animal Learning & Behavior, 26, 416-426. Andersson, M. (1982). Female choice for extreme tail length in a widowbird. Nature, 299, 818-820. Assenmacher, I., & Jallegeas, M. (1980). In A. Epple & M. H. Stetson (Eds.), Avian endocrinology: Vol. 3 (pp. 183-286). New York, NY: Academic Press. Beecher, M., & Beecher, I. (1979). Sociobiology of bank swallows: Reproductive strategy of the male. Science, 205, 1282-1285.Bennett, A. T. D., Cuthill, I. C., Partridge, J. C., & Maier, E. J. (1996). Ultraviolet vision and mate choice in zebra finch. Nature, 380, 433-435. Bennett, A. T. D., Cuthill, I. C., Partridge, J. C., & Lunau, K. (1997). Ultraviolet plumage colors predict mate preferences in starlings. Proceedings of the National Academy of Science, USA, 94, 8618-8621. Benoit, J. (1938). R�le des yeux et de la voie nerveuse oculo-hypophysaire dans la gonadostimulation par la lumi�re arteficiell chez le Canard domestique. Comptes Rendus des Seances. Societe de Biologie (Paris), 129, 231-234. Benoit, J., & Assenmacher, I. (1966). Research on the photosensitivity of superficial and deep neural receptors in gonadal stimulation by visible radiation in the immature Pekin duck. Comptes Rendus Hebdomadaires des Seances de l Academie des Sciences - D: Sciences Naturelles, 262, 2750-2752. Birkhead, T. (1979). Mate guarding in the magpie, Pica pica. Animal Behaviour, 27, 866-874. Bluhm, C. K., Phillips, R. E., Burke, R. E., & Gupta, G. N. (1983). Effects of male courtship and gonadal steroids on pair formation, egg-laying, and serum LH in canvasback ducks (Aythya valisinaria). Journal of Zoology (London), 204, 185-200. Bons, N. (1976). Retinohypothalamic pathway in the duck (Anas platyrhynchos). Cell & Tissue Research, 168, 343-360. Bons, N., Jallageas, M., & Assenmacher, I. (1975). Influence des recepteurs retiniens et extra-retiniens dans la stimulation testiculaire de la Caille par les "jours longs". Journal de Physiologie. (Paris), 71, 265-266. Carbaugh, B. T., Schein, M. W., & Hale, E. B. (1962). Effects of morphological variations of chicken models of sexual responses of cocks. Animal Behavior, 10, 235-238. Cooper, W. T., & Forshaw, J. M. (1979). The birds of paradise and bower birds. Boston, MA: D. R. Godine. Cooper, M. L., Pickard, G. E., & Silver, R. (1983). Retino-hypothalamic pathway in the dove demonstrated by anterograde HRP. Brain Research Bulletin, 10, 715-718. Crawford, L. L., & Akins, C. K. (1993). Stimulus control of copulatory behavior in male Japanese quail. Poultry Science, 72, 722-727. Cusato, B., & Domjan, M. (1998). Special efficacy of sexual conditioned stimuli that include species typical cues: Tests with a conditioned stimuli preexposure design. Learning & Motivation, 29, 152-167. Domjan, M. (1993). The principles of learning and behavior. Pacific Grove, CA: Brooks/Cole Publishing Company. Domjan, M. (1994). Formulation of a behavior system for sexual conditioning. Psychonomic Bulletin & Review, 1, 421-428. Domjan, M., Blebois, E., & Williams, J. (1998). The adaptive significance of sexual conditioning: Pavlovian control of sperm release. Psychological Science, 9, 411-415. Domjan, M., Akins, C. K., & Vandergriff, D. H. (1992). Increased responding to female stimuli as a result of sexual experience: Tests of mechanisms of learning. Quarterly Journal of Experimental Psychology, 45B, 139-157. Domjan, M., & Hall, S. (1986). Sexual dimorphism in the social proximity behavior of Japanese quail. Journal of Comparative Psychology, 100, 68-71. Domjan, M., Greene, P., & North, N. C. (1989). Contextual conditioning and the control of copulatory behavior by species-typical sign stimuli in male Japanese quail. Journal of Experimental Psychology: Animal Behavior Processes, 15, 147-153. Domjan, M., Lyons, R., North, N. C., & Bruell, J. (1986). Sexual Pavlovian conditioned approach behavior in male Japanese quail (Coturnix coturnix japonica). Journal of Comparative Psychology, 100, 413-421. Domjan, M., & Nash, S. (1988). Stimulus control of social behavior in male Japanese quail (Coturnix coturnix japonica). Animal Behavior, 36, 1006-1015. Domjan, M., O�Vary, D., & Greene, P. (1988). Conditioning of appetitive and consummatory sexual behavior in male Japanese quail. Journal of Experimental Analysis of Behavior, 50, 505-519. Donham, R. S. (1979). The annual cycle of plasma leutinizing hormone and sex hormones in male and female mallards (Anas platyrhynchos). General & Comparative Endocrinology, 29, 152-155. Ebihara, S., & Kawamura, H. (1981). The role of the pineal organ and the suprachiasmatic nucleus in the control of circadian locomotor rhythms in the Java sparrow, Padda oryzivora. Journal of Comparative Physiology, 1412, 207-214. Eiserer, L. A., & Swope, R. L. (1980). Acquisition of behavioral control by static visual features of an imprinting object: Species Generality. Animal Learning & Behavior, 8, 481-484. Fisher, A. E., & Hale, E. B. (1957). Stimulus determinants of sexual and aggressive behaviour in male domestic fowl. Behaviour, 10, 309-323. Gillard, E. T. (1969). Birds of paradise and bower birds. Garden City, N.Y.: Natural History Press. Hartwig, H. G. (1974). Electron microscopic evidence for a retinohypothalamic projection to the suprachiasmatic nucleus for Passer domesticus. Cell & Tissue Research, 153, 89-99. Hollis, K. L. (1997). Contemporary research on Pavlovian conditioning: A "new" functional analysis. American Psychologist, 52(9), 956-965. Hollis, K. L., Cadieux, E. L., & Colbert, M. M. (1989). The biological function of Pavlovian conditioning: A mechanism for mating success in the blue gourami (Trichogaster trichopterus). Journal of Comparative Psychology, 103, 115-121.Hollis, K. L., Pharr, V. L., Dumas, M. J., Britton, G. B., & Field, J. (1997). Classical conditioning provides paternity advantage for territorial male blue gouramis (Trichogaster trichopterus). Journal of Comparative Psychology, 111(3), 219-225. Johnsgard, P. A. (1994). Arena birds. Washington, D.C. Smithsonian Institution Press. Lehrman, D. S., & Wortis, R. (1960). Previous breeding experience and hormone-induced incubation behavior in the ring dove. Science, 132, 1667-1668. Lumpkin, S., Kessel, K., Zenone, P.G., & Erickson, C.J. (1982). Proximity between the sexes in ring doves: Social bonds or surveillance. Animal Behaviour, 30, 506-513.Mattocks Jr., P. W., Farner, D. S., Follett, B. K. (1976). The annual cycle in leutinizing hormone in the plasma of intact and castrated white-crowned sparrows (Zonotrichia leucophrys gambelii). General & Comparative Endocrinology, 30, 156-161. Meier, R. (1973). Autoradiographic evidence for a direct retino-hypothalamic projection in the avian brain. Brain Research, 53, 417-421. Menaker, M., Roberts, R., Elliot, J., & Underwood, H. (1970). Extraretinal light perception in the sparrow, Vol III: The eyes do not participate in photoperiodic photoreception. Proceedings of the National Academy of Sciences, Washington, 67, 320-325. Nash, S., Domjan, M., & Askins, M. (1989). Sexual discrimination learning in male Japanese quail (Coturnix coturnix japonica). Journal of Comparative Psychology, 103, 347-358. Papini, M. R., & Bitterman, M. E. (1990). The role of contingency in classical conditioning. Psychological Review, 97, 396-403. Petrie, M. Halliday, T., & Sanders, C. (1991). Peahen prefer peacocks with elaborate trains. Animal Behaviour, 41, 323-331. Rands, M. R. W., Ridley, M. W., & Lelliott, A.D. (1984). The social organization of feral peafowl. Animal Behaviour, 32, 830-835. Reperant, J. (1973). Nouvelles donness sur les projections visuelles chez le Pigeon (Columba livia). Journal fuer Hirnforschung, 14, 307-312. Rescorla, R. A. (1967). Pavlovian conditioning and its proper control procedures. Psychological Review, 74, 71-80. Rowan, W. (1925). Relation of light to bird migrations and developmental changes. Nature (London), 115, 494-495. Scanes, C. G. (1974). Some in vitro effects of synthetic thyrotrophin releasing factor on the secretion of thyroid stimulating hormone from the anterior pituitary gland of the domestic fowl. Neuroendocrinology, 15, 1-9. Schein, M. W., & Hale, E. B. (1957). The head as a sexual stimulus for orientation and arousal of sexual behavior of male turkeys. Anatomical Record, 128, 617-618. Schein, M. W., & Hale, E. B. (1965). Stimuli eliciting sexual behavior. In F. A. Beach (Ed.), Sex and behavior (pp. 440-482). New York, NY: Wiley. Schoettle, H. E. T., & Schein, M. W. (1959). Sexual reactions of male turkeys to deviations from a normal female head model. Anatomical Record, 134, 635. Sharp, P. J., & Moss, R. (1981). A comparison of the response of captive willow ptarmigan (Lagopus lagopus lagopus), red grouse (L. l. scoticus) and hybrids to increasing day length with observations on the modifying effects of nutrition and crowding in red grouse. General & Comparative Endocrinology, 45, 181-188. Shields, K. M., Yamamoto, J. T., & Millam, J. R. (1989). Reproductive behavior and LH levels of cockatiels (Nymphicus hollandicus) associated with photostimulation, nest-box presentation, and degree of mate access. Hormones & Behavior, 23, 68-82. Stokkan, K. A., & Sharp, P. J. (1980). Seasonal changes in the concentrations of plasma leutinizing hormone and testosterone in willow ptarmigan (Lagopus lagopus lagopus) with observations on the effects of permanent short days. General & Comparative Endocrinology, 40, 109-115. Timberlake, W. (1993). Behavior systems and reinforcement: An integrative approach. Journal of Experimental Analysis of Behavior, 60, 105-128. Timberlake, W. (1994). Behavior systems associationism, and Pavlovian conditioning. Psychonomic Bulletin & Review, 1, 405-420. Timberlake, W., & Lucas, G. A. (1989). Behavior systems and learning: From misbehavior to general principles. In S. B. Klein & R. R. Mowrer (Eds.), Contemporary learning theories: Instrumental conditioning theory and the impact of biological constraints on learning (pp. 237-275). Hillsdale, NJ: Erlbaum. Twitmeyer, E. B. (1974). A study of the knee jerk. Journal of Experimental Psychology, 103, 1047-1066.Wilson, F. E., & Follet, B. K. (1974). Plasma and pituitary leutinizing hormone in intact and castrated tree sparrows (Spizella arborea) during a photoinduced gonadal cycle. General & Comparative Endocrinology, 23, 82-93. Wilson, R. H. (1974). Agonistic postures and latency to the first interaction during initial pair encounters in red jungle fowl (Gallus gallus). Animal Behaviour, 22, 75-82. Wingfield J. C. (1980) Fine temporal adjustment of reproductive functions. In A. Epple & M. H. Stetson (Eds.) Avian endocrinology (pp. 149-166). Wingfield, J. C., & Farner, D. S. (1993). Endocrinology of Reproduction in Wild Species. In D. S. Farner, J R. King, & K. C. Parkes, (Eds.), Avian biology: Volume IX (pp. 163-327). San Diego, CA: Academic Press. Yokoyama, K., & Farner, D. S. (1978). Induction of Zugunruhe by
photostimulation of encephalic receptors in white-crowned sparrows. Science,
201, 76-79. For a good example of feather display, see this movie about Peafowl. A species in trouble: the Lesser Prairie Chicken. Viewing of the Lesser Prairie Chicken. Information about Temmick's Tragopan Pheasants. Click here for information about Bower Birds. Click Here for a list of photographers who's work has been added to this chapter.
|