|
|
|
The literature
provides only moderate evidence of a role for anticipation in mediation of short-term
retention in pigeons. The strongest evidence comes from a spatial task in which
pigeons appear able to remember locations not yet chosen. The differential outcome
literature is consistent with mediation by anticipated affective outcomes, but may reflect
instead persisting emotional or behavioral reactions elicited by samples correlated with
outcomes that differ affectively. Research using many-to-one sample-to-choice
mapping procedures is similarly not definitive with regard to whether the codes mediating
retention have anticipatory content. Moreover, research has failed to obtain
evidence that short-term retention can be mediated by either (a) anticipated outcomes
which are affectively equivalent or (b) simple response intentions. Finally,
although analysis of confusion errors and interference effects suggests prospective coding
in choice matching with lines or colors (at least in procedures involving the simultaneous
presentation of three choice stimuli), the small sample size and mixed statistical
evidence suggests caution in evaluating the theoretical significance of those
results. Continued research is necessary to clarify the role of anticipation as a
mediator of short-term retention in pigeons.
|

I. Introduction
The reader may view as a truism the notion that at
least some species of animals are capable of anticipating future events. Such
anticipation, one might suppose, could be triggered by the occurrence of particular
stimuli that in the past have been followed reliably by some event of significance (e.g.,
the occurrence of food). Similarly, one might suppose further, such anticipation
could be evoked by the actions in which an animal is presently engaged to the extent that
those actions have in the past been followed reliably by some event of significance.
Pavlovian or classical conditioning experiments represent a laboratory instantiation of
situations in which stimuli are established as reliable predictors of other stimuli.
Instrumental or operant conditioning experiments represent a laboratory instantiation of
situations in which actions are reliably followed by stimulus consequences.
Although it may appear obvious that Pavlovian and
instrumental learning tasks would result in organisms learning to anticipate future events
on the basis of current stimuli or present actions, respectively, the notion that animals
can anticipate future events was strongly resisted by the majority of learning theorists
from the early 1900s to the late 1960s. These theorists, often referred to as S-R
(stimulus-response) behaviorists, prominent among whom were Thorndike, Watson, Guthrie,
Hull, and Spence, expended considerable effort in developing explanations of behavior that
eschewed reference to anticipation or expectancy. The S-R behaviorists argued that
an adequate explanation of the behavior of animals, including humans, could be provided
without introducing cognitive processes such as anticipations and expectancies.
Instead, they argued, behavior could be explained in terms of simpler psychological
processes, prominent among which was the process of new stimulus-response habit or reflex
formation. On this view, learning involves the strengthening and weakening of
stimulus-response habits or reflexes as a result of the organism's experience.
For example, Thorndike (1898) proposed the well
known Law of Effect which maintained that an action producing a favorable outcome
became more strongly connected to the stimuli present when that action
occurred. Similarly, an action producing an unfavorable outcome became less strongly
connected to the stimuli present when that action occurred. Hence, the organism's
behavior adjusted to the situation through this mechanical (unthinking, automatic) process
of strengthening and weakening of stimulus-response habits as a function of
experience. That is, as an organism's experience in a situation increased, the
stimuli present would, with increasing probability, trigger reactions that in the past had
produced a favorable outcome. Given that an adequate explanation of behavioral
adjustment as a function of experience could be provided in terms of the low-level
psychological process of the strengthening and weakening of stimulus-response habits, the
S-R behaviorists contended that the introduction of psychologically more complex
mechanisms, such as expectations, was superfluous.
However, some theorists in the pre 1970 era, most
notably Tolman and his associates, argued that an adequate explanation of behavioral
adjustment could not be based on a law-of-effect process. In
particular, Tolman (1932, 1948) argued that organisms acquire knowledge about an
environment as a function of exposure to that environment. Hence, Tolman maintained
that organisms do learn "what-leads-to-what". In a maze situation, for
example, Tolman argued that organisms do not learn a series of stimulus-response habits
which get them from the starting point to the goal location, as the S-R behaviorsts
maintained. Instead, Tolman contended, organisms learn about the geographical
properties of the maze. That is, organisms form an internal representation of the
maze, a "cognitive map", and learn to expect food to be encountered in the
location where it had been encountered previously.
Much of Tolman's research was devoted to
demonstrating the inadequacies of a law-of-effect based stimulus-response explanation of
behavior. In so doing, Tolman hoped to convince other theorists that the
introduction of higher-level psychological processes, such as expectations and cognitive
maps, was justified - - indeed, necessary - - to explain behavioral adjustment. One
aspect of Tolman's research involved demonstrating that learning could occur even when
responses were not followed by favorable or unfavorable outcomes. For example,
Tolman and Honzik (1930b) provided evidence that rats learn the layout of a complex maze
even when no favorable outcome was present in the goal box (i.e., the goalbox was empty),
learning that clearly could not be explained by a law-of-effect process. Another
aspect of Tolman's research involved demonstrating that rats represent the geographical
properties of mazes and learn to expect food in a particular location in that
environment. For example, Tolman and Honzik (1930a) and Tolman, Ritchie, and Kalish
(1946a, b) provided evidence that rats learn where the goal is located, rather than the
specific responses required to get to the goal.
Although the research of Tolman and his associates
clearly revealed the inadequacy of explanations of behavior based solely on the Law of
Effect, that research did not convince the majority of theorists that the introduction of
expectancies and cognitive maps as explanatory constructs was required. Indeed, Hull
(1943, 1952), Spence (1956, 1960), and others (often referred to as neo S-R behaviorists)
sought adequate explanations of the phenomena discovered by Tolman and his associates
without abandoning a strict stimulus-response analysis of behavior. Although the neo
S-R behaviorists introduced explanatory mechanisms in addition to the Law of Effect, they
maintained the fundamental proposition that learning involved nothing more than the
strengthening and weakening of stimulus-response habits. However, research in
Pavlovian conditioning conducted in the late 1960s challenged this fundamental
proposition.
In particular, Kamin's (1968, 1969) discovery of
blocking, Rescorla's (1966, 1968) work on contingency effects, and Wagner's (1969; Wagner,
Logan, Haberlandt & Price, 1968) work on relative cue validity effects made clear the
inadequacy of a stimulus-response conception of Pavlovian conditioning. This
research revealed that learning occurred in Pavlovian conditioning only when a stimulus
was informative with regard to the probability of occurrence of a second stimulus. A
consideration of the details of this research is beyond the scope of the present article,
and it sufficient to note that the work of these researchers revealed that organisms in
Pavlovian conditioning situations learn something in addition to, or, more likely, other
than stimulus-response habits. Although these data suggest that the "something
else" which is learned is an expectancy for the occurrence or nonoccurrence of the
second stimulus upon presentation of the first, they do not compel such a conclusion.
More contemporary research in both Pavlovian and
instrumental conditioning has, however, produced definitive evidence that organisms do
indeed learn "what-leads-to-what"; that is, that organisms do learn to
anticipate the consequence of stimuli and/or their own actions. Particularly
definitive evidence for anticipation has been provided by studies in which the value of
the consequent has been modified after conditioning. In these studies, initially a
stimulus (e.g., Holland & Rescorla, 1975; Holland & Straub, 1979) or an action
(e.g., Colwill & Rescorla, 1985; Dickinson, Campos, Varga, & Balleine, 1996) has
been established as a reliable predictor of an outcome (e.g., sucrose pellets or standard
food pellets). Subsequently, the value of the consequent has been reduced (e.g., by
satiating the animal on that consequent, or by making the consequent aversive by pairing
it with induced illness). During testing in which the consequent is no longer
presented, animals react to the predictor, whether stimulus or action, in a way clearly
revealing that they had learned what is predicted by that stimulus or action. For
example, rats that had initially reacted with agitated excitement to a tone paired with
sucrose, show little excitement to the tone following devaluation of the sucrose.
Similarly, rats that had earlier eagerly pressed a lever to obtain sucrose pellets, will,
after devaluation of sucrose, be reluctant to press the lever.
The evidence is now sufficient to convince the
majority of learning theorists that some species of animals are capable of anticipating
the consequences of environmental stimuli and/or their own actions. The authors of
this article are memory researchers, and hence are concerned primarily with how learned
information is utilized subsequent to its acquisition (Spear & Riccio, 1994). In
particular, our interest is focussed on the mechanisms and processes which permit
organisms to retain information over relatively short intervals (i.e., seconds to minutes)
and to utilize that information at the end of the interval. One abiding interest in
our laboratory has been whether animals, specifically pigeons, learn to anticipate future
events and whether such anticipation can also mediate retention or memory-based
performance. The remainder of this article is concerned with this question.
In the next section (II), we describe a procedure,
known as delayed matching-to-sample, which has been particularly useful in analytical
investigations of short-term retention in pigeons. In the following section (III),
we distinguish between retrospective and prospective coding (the latter, but not the
former, involving anticipation of future events). In that section, we ask whether
there is convincing evidence that pigeons, at least under some circumstances, code
prospectively (and, hence, reveal evidence for mediation of retention by anticipated
future events). We then turn (section IV) to a consideration of whether an
anticipated outcome (e.g., food or water for correct responding) can mediate
retention. In section V, we consider the issue of whether pigeons can use a
"simple" response anticipation (i.e., to respond or not to respond) to guide or
mediate short-term retention. And, finally, in the last section (VI) we summarize
the preceding discussion and address some questions and issues raised by this work, and
consider the implications of those questions and issues for our understanding of memory
processes and mechanisms.
II. The Delayed
Matching-To-Sample Procedure
The matching-to-sample procedure has been used in
numerous investigations of short-term retention in pigeons. Matching is a
conditional discrimination task because the reinforcement contingencies vary from trial to
trial. For example, trials might always involve a choice between pecking a key
illuminated with red light or pecking a key illuminated with green light. These
choice stimuli are typically presented on the outer-most keys in a row of three, and their
positions are counterbalanced such that, over trials, each stimulus appears equally often
on the right and left keys. On some trials, pecking the red key produces access to
food (and hence pecking the green key does not), and on other trials the reverse is the
case (i.e., pecking the green key produces access to food and pecking the red key does
not). A conditional cue, typically referred to as the sample stimulus, is presented
at the beginning of each trial and indicates the reinforcement contingencies in effect on
that trial. The sample stimulus is typically projected on the center key and, in the
identity version of matching, is physically identical to the choice stimulus that must be
pecked to produce reinforcement on that trial. Hence, in the present example, a red
sample stimulus indicates that pecking the red, but not the green, choice stimulus will
produce reinforcement, whereas a green sample stimulus indicates that pecking the green,
but not the red, choice stimulus will produce reinforcement. Which sample  stimulus is presented (and hence which choice stimulus is correct) varies
randomly from trial to trial. In the simultaneous version of matching, the sample
stimulus remains available on the center key until the animal responds to one of the
choice stimuli (click here for animated
illustration). stimulus is presented (and hence which choice stimulus is correct) varies
randomly from trial to trial. In the simultaneous version of matching, the sample
stimulus remains available on the center key until the animal responds to one of the
choice stimuli (click here for animated
illustration).
After 10 to 15
sessions of training, with each session comprised of 48 trials, most pigeons peck the
matching (and hence correct) choice stimulus on 95% or more of the trials. Because
in simultaneous matching the sample stimulus remains available until the choice response
has been made, accurate performance requires only that the organism learn the rules
relating sample stimulus and correct choice stimulus (e.g., choose green if green sample,
and choose red if red sample). An additional short-term retention requirement can be
imposed by temporally separating presentation of the sample stimulus and presentation of
the choice stimuli. This is accomplished by interpolating a short delay or retention
interval, typically a few seconds in duration, between termination of the sample stimulus
and presentation of the choice stimuli. Because
the sample is not present at the time the choice response is emitted in the delayed
version of matching-to-sample, accurate choice requires utilization of information derived
during sample presentation and retained during the delay interval (click here for animated illustration).
III. Retrospective and Prospective Coding
The Distinction
As noted in the preceding section,
accurate performance in delayed matching-to-sample requires the retention and utilization
of information derived during sample presentation. In the early 1980s, several
investigators recognized that the information or code which mediates short-term retention
in delayed matching-to-sample could have either a retrospective or prospective content
(e.g., Grant, 1981; Honig & Thompson, 1982; Riley, Cook, & Lamb, 1981; Roitblat,
1980). In particular, the animal could retain features of the sample stimulus during
the delay, a retrospective or "backward-looking" code. During testing, the
retrospective code, in combination with the rules learned during simultaneous matching,
would be sufficient to generate accurate choice performance. Alternatively, the
rules learned during simultaneous matching could be activated during sample presentation
to generate an anticipatory code. That is, the code could represent features of the
correct choice stimulus, a prospective or "forward-looking" code.
The distinction between
retrospective and prospective (or anticipatory) code content is most readily illustrated
in the symbolic (or arbitrary) version of delayed matching. As opposed to identity
matching, in which the correct choice stimulus is physically identical to the sample
presented on that trial, in arbitrary matching the relation between sample and correct
choice stimulus is established arbitrarily by the experimenter. For example, as
illustrated in the accompanying animation, the samples might be red and green colors and
the choice stimuli might be white bars oriented vertically or
horizontally: And, for example, vertical might be correct (and hence pecking it is
reinforced) on red-sample trials, and horizontal might be correct (and hence pecking it is
reinforced) on green-sample trials (click
here for animated illustration).
In this example, the code could have
a retrospective content in that the code preserves the visual features of the
sample
stimulus (i.e., "redness" or "greenness" on any particular trial). If the code is active at the time of
testing, correct responding could be generated by the learned rules of the matching
procedure (i.e., "if red, peck vertical" and "if green, peck
horizontal"), (click here for
animated illustration).
Alternatively, the code could have
an anticipatory or prospective content in that the code preserves the visual features of
the correct choice stimulus (i.e., "vertical" or "horizontal" on any
particular trial). In this case, the learned rules of the matching procedure are invoked during the sample presentation period to generate
the anticipatory code. At the time of testing, the active code is sufficient to
generate correct responding (click here
for animated illustration).
Evidence for Prospective Code Content
In the present article, the primary
question of interest is whether there is evidence that pigeons employ anticipatory or
prospective codes in delayed matching (and in related procedures that require information
to be maintained over relatively short intervals). In this section we consider
evidence for prospective coding obtained in standard and many-to-one matching procedures,
and from a procedure requiring retention of spatial information over short
intervals.
In standard matching. We use the term standard
matching to refer to identity or arbitrary matching procedures in which (a) the
samples are colors, lines and/or forms and (b) each sample stimulus is associated with a
unique choice stimulus. Experiments employing the standard procedure have provided
some evidence for prospective code content. Perhaps the most powerful of this
evidence was reported by Roitblat (1980, Exp. 3). Roitblat's procedure was based on
an experiment by Conrad (1964) in which human subjects were employed.
Conrad (1964) was interested in
whether humans would code visually presented letters in terms of their visual properties
(e.g., letter B as the visual image of B) or in terms of their acoustic properties (e.g.,
letter B as the sound of B). He argued that if subjects code letters visually, then
memory errors should reflect the visual similarity between letters (e.g., misremembering B
as visually similar R). If subjects code letters acoustically, then memory errors
should reflect the acoustic similarity between letters (e.g., misremembering B as
acoustically similar V). Conrad found that errors were related to acoustic, rather
than visual, similarity, suggesting that the letters were coded in terms of their acoustic
properties.
In the case of pigeons performing
delayed matching with colors or line orientations, it is reasonable to assume that the
code represents visual features of the stimuli. Hence, the question of interest is
not the modality of the code (e.g., visual or acoustic), but rather whether the code
preserves visual features of the sample (i.e., retrospective content) or visual features
of the correct choice stimulus (i.e., prospective content). As Roitblat (1980)
noted, Conrad's (1964) research suggests a way to determine whether pigeons in delayed
matching code features of the sample or code features of the correct choice
stimulus. Specifically, if pigeons code retrospectively, then memory errors should
reflect similarity among the samples (e.g., confusing similar orange and red samples
rather than dissimilar orange and blue samples). If, on the other hand, pigeons code
prospectively, then memory errors should reflect similarity among the choice stimuli
(e.g., confusing similar vertical and nearly vertical choice stimuli rather than
dissimilar vertical and horizontal choice stimuli).
To implement this test, Roitblat
(1980) trained birds in an arbitrary matching task involving three samples and three
choice stimuli. Two birds were trained with orange, red, and blue color samples and
vertical, nearly vertical, and horizontal line  choice stimuli, and the remaining bird
received lines as samples and colors as choice stimuli. As shown in Figure 1, similar pairs of samples were associated with dissimilar pairs
of choice stimuli, and dissimilar pairs of samples were associated with similar pairs of
choice stimuli. A delay ranging between .25 and 5.60 s intervened between sample
termination and presentation of all three choice stimuli.
choice stimuli, and the remaining bird
received lines as samples and colors as choice stimuli. As shown in Figure 1, similar pairs of samples were associated with dissimilar pairs
of choice stimuli, and dissimilar pairs of samples were associated with similar pairs of
choice stimuli. A delay ranging between .25 and 5.60 s intervened between sample
termination and presentation of all three choice stimuli.
To further illustrate the rationale
underlying Roitblat's (1980) experiment, consider a trial involving the orange sample and
the nearly vertical correct choice. On this trial type, the birds will err by
choosing either the vertical line associated with the blue sample or the horizontal line
associated with the red sample. If the birds are remembering "orange"
(retrospective code), they should tend to confuse orange with red and should therefore
select the horizontal line. If the birds are remembering "nearly vertical
line" (prospective code), they should tend to confuse nearly vertical with vertical
and should therefore select the vertical line.
The data revealed that confusion
between similar choice stimuli increased faster across the retention interval than did
confusion between similar samples, although the effect was statistically significant only
for the birds receiving color samples and line choice stimuli. This finding provides
evidence for prospective code content, and hence suggests that the anticipation of a color
or a line can mediate retention in delayed matching. Roitblat and Scopatz (1983)
maintained that this conclusion was further strengthened by an analysis of proactive
interference. Roitblat and Scopatz employed three pigeons and an identity-matching
task with red, green and blue stimuli. As in Roitblat's (1980) experiment, following
sample presentation all three stimuli were presented simultaneously for a choice.
They examined choice on each trial (after the first) as a function of the sample presented
and choice made on the immediately preceding trial. They found that choices on a
trial were influenced by the choice made, rather than by the sample presented, on the
preceding trial. Roitblat and Scopatz argued that this result also suggests that
pigeons remember "what to do" rather than "what happened" in standard
delayed matching (i.e., a prospective code).
It may be, however, that the
Roitblat and Scopatz (1983) finding can be explained without assuming that samples are
coded prospectively. It could be argued, for example, that pecking a choice stimulus
activates a representation of that stimulus. Moreover, the representation of the
chosen stimulus might interfere with the retrospective code of the sample presented
earlier on that trial (retroactive interference effect). It then follows that, at
the time of choice on the succeeding trial, the animal is more likely to remember the
stimulus chosen than the sample presented on the preceding trial. This persisting
memory for the stimulus chosen on the preceding trial would compete with memory for the
sample on the current trial for control over choice responding. Hence, choice on a
trial should be related more strongly to the stimulus chosen than to the sample presented
on the preceding trial. It is also possible that sample coding errors contributed to
Roitblat and Scopatz's finding. Again assume that pigeons code samples
retrospectively. Assume further that errors in delayed matching often reflect the
misidentification of the sample. For example, a red sample activates the code
"green". On such a trial, the code "green" would produce choice
of the green test stimulus. If memory of either (or both) the code activated by the
sample (i.e., "green") or the stimulus chosen (i.e., green) persisted until
choice on the succeeding trial, then choice on that trial should be related to the
stimulus chosen (i.e., green) rather than the sample presented (i.e., red) on the
preceding trial. In our view, therefore, one could maintain the view that samples
are coded retrospectively and provide an adequate explanation of Roitblat and Scopatz's
data. If so, then their data do not provide definitive evidence for prospective
coding.
It should be noted that Urcuioli and
Zentall (1986) obtained data which led them to conclude that pigeons code retrospectively
in standard delayed matching. They employed highly discriminable colors (red and
green) and less discriminable lines (horizontal and vertical) as stimuli. Some birds
received color samples mapped to color or line choice stimuli. Other birds received
line samples mapped to color or line choice stimuli.
Urcuioli and Zentall (1986) reasoned
that retention should be a function of the discriminability of the event being retained;
that is, highly discriminable events (colors in the present case) should be remembered
more effectively than less discriminable events (lines in the present case). Hence,
if pigeons code retrospectively, then birds receiving highly discriminable colors as
samples should demonstrate better retention than those receiving less discriminable lines,
regardless of the dimension of the choice stimuli. If, on the other hand, pigeons
code prospectively, then birds receiving highly discriminable colors as choice stimuli
should demonstrate better retention than those receiving less discriminable lines,
regardless of the dimension of the sample stimuli.
After training, pigeons received two
retention tests, one with delays ranging from 0 to 4 s, and a second with delays ranging
from 0 to 8 s. Urcuioli and Zentall (1986) found that sample dimension influenced
rate of forgetting, in that retention functions were less steep for pigeons receiving
color samples than for those receiving line samples. In contrast, choice dimension
had little effect on rate of forgetting, although accuracy collapsed across delays was
higher in pigeons trained with color choice stimuli than in those trained with line choice
stimuli. Urcuioli and Zentall argued that these results suggest that the pigeons
coded the samples retrospectively and hence demonstrated better retention when the samples
were the more discriminable colors than when they were the less discriminable lines.
However, Urcuioli and Zentall's
(1986) conclusion is open to criticism on both conceptual and interpretational
grounds. At a conceptual level, Roitblat (1993, p. 180) has argued that both
prospective and retrospective coding accounts predict that, as found by Urcuioli and
Zentall, more difficult to discriminate samples would result in lower accuracy and more
rapid forgetting. Roitblat notes that samples must be identified before a
prospective code can be activated, and that more difficult to discriminate stimuli are
likely to be identified less accurately and with less strength. It then follows that
more difficult to discriminate samples should result in reduced accuracy and faster
forgetting than samples which are more easily discriminated, even if birds code
prospectively.
At a methodological level, it is
unclear whether the significant Group x Delay interaction obtained by Urcuioli and Zentall
(1986) reflects between-group differences in rate of forgetting or a ceiling
artifact. With regard to the latter, it should be noted that performance on 0-s
delay trials was close to 100% in groups trained with color samples (estimating from
Figures 1 through 4 in their article, accuracy appears to range from 96 to 98% correct at
the 0-s delay). Hence, it could be that accuracy differences as a function of sample
dimension were obscured by a ceiling artifact at the 0-s delay. At increasingly
longer delays, performance dropped below ceiling, allowing between-group differences to
emerge. It may be, therefore, that sample dimension did not influence rate of
forgetting, in spite of the significant Group x Delay interaction.
Some support for this view is
provided by examining performance only at longer delays, where the operation of a ceiling
artifact is less likely (see Figure 2;
the data  points in the left
panel were estimated from Figure 2, and those in the right panel from Figure 4, in
Urcuioli & Zentall, 1986). With either line or color choice stimuli, whether one
examines performance at 2- and 4-s delays in the short-delay test (not shown in Figure 2),
or performance at 2-, 4-, and 8-s delays in the long-delay test, there is no evidence that
forgetting is more rapid on line-sample trials than on color-sample trials. Indeed,
with color choice stimuli (Figure 2
right panel), rate of forgetting appears to be somewhat faster on color- than on
line-sample trials. Although accuracy collapsed across delay length was higher with
color than with line samples, this result is easily explained in terms of the differential
effectiveness with which easy versus difficult to discriminate samples activate the
correct prospective code. points in the left
panel were estimated from Figure 2, and those in the right panel from Figure 4, in
Urcuioli & Zentall, 1986). With either line or color choice stimuli, whether one
examines performance at 2- and 4-s delays in the short-delay test (not shown in Figure 2),
or performance at 2-, 4-, and 8-s delays in the long-delay test, there is no evidence that
forgetting is more rapid on line-sample trials than on color-sample trials. Indeed,
with color choice stimuli (Figure 2
right panel), rate of forgetting appears to be somewhat faster on color- than on
line-sample trials. Although accuracy collapsed across delay length was higher with
color than with line samples, this result is easily explained in terms of the differential
effectiveness with which easy versus difficult to discriminate samples activate the
correct prospective code.
In many-to-one matching. As
noted previously in this article, matching procedures can be differentiated on the basis
of whether the matching relation is based on physical identity or is established
arbitrarily by the experimenter. A second distinction between matching procedures is
based on the sample-to-choice mapping ratio. In the procedures considered so far,
that ratio has been one-to-one: each sample stimulus is associated with one and only one
correct choice stimulus. Hence, the number of stimuli in the sample set is
equivalent to the number of stimuli in the choice set. Therefore, the number of
codes required to mediate accurate retention in a one-to-one procedure is independent of
whether the codes represent features of the sample stimuli (retrospective coding) or the
correct choice stimuli (prospective coding).
Alternatively, in a many-to-one
sample-to-choice mapping, two or more sample stimuli are associated with each choice
stimulus (e.g., Grant, 1982a, 1982b, 1991; Grant & Spetch, 1993; Santi & Roberts,
1985; Urcuioli, DeMarse, & Zentall, 1994; Zentall, Jagielo, Jackson-Smith, &
Urcuioli, 1987; Zentall, Urcuioli, Jagielo, & Jackson-Smith, 1989). A
many-to-one procedure appears particularly conducive to prospective coding because such
coding could reduce the number of codes required for accurate performance. For
example, consider a procedure in which three samples are associated with one choice
stimulus, and three different samples are associated with a second choice stimulus (a 3:1
many-to-one mapping). Remembering the sample presented on each trial would require
six different codes (one for each of the six different sample stimuli).
Alternatively, remembering the correct choice stimulus on each trial would require only
two different codes (one for each of the two different choice stimuli).
Urcuioli, Zentall, Jackson-Smith,
and Steirn (1989, Exp. 2), trained birds in a many-to-one matching procedure with color
(red and green) and line (vertical and horizontal) sample stimuli. All trials
involved a choice between vertical and horizontal lines presented on the outer keys.
Vertical was the correct choice on trials involving a red or vertical sample, and  horizontal was the correct choice on trials
involving a green or horizontal sample (see top panel in Figure 3). Hence, the red and
vertical samples were functionally equivalent (i.e., both were signals to choose vertical)
and the green and horizontal samples were functionally equivalent (i.e., both were signals
to choose horizontal). horizontal was the correct choice on trials
involving a green or horizontal sample (see top panel in Figure 3). Hence, the red and
vertical samples were functionally equivalent (i.e., both were signals to choose vertical)
and the green and horizontal samples were functionally equivalent (i.e., both were signals
to choose horizontal).
If the animal coded prospectively,
only two different codes would be required. Specifically, the two samples associated
with the vertical choice stimulus would activate the code "vertical" (or
"peck vertical") and the two samples associated with the horizontal choice
stimulus would activate the code "horizontal" (or "peck
horizontal"). If, on the other hand, the animal coded retrospectively, four
different codes (one to represent each of the four samples) would be required.
Hence, a many-to-one mapping appears particularly suited to use of prospective coding in
which each of the two functionally equivalent samples activates the same code.
Urcuioli et al. (1989, Exp. 2) did
obtain evidence that functionally equivalent samples activate the same code. They
reasoned that if functionally equivalent samples activate the same code, then one might
observe transfer of performance between functionally equivalent samples. To assess
this possibility, the many-to-one training phase described previously was followed by a
second training phase in which the color samples employed in the many-to-one training
phase were associated with new choice stimuli, circle and dots (see middle panel in
Figure 3). Following acquisition, the
animals received a transfer test in which the line samples from many-to-one training were
followed by the circle and dots choice stimuli from one-to-one training (see bottom panel
in Figure 3).
If many-to-one training resulted in a
color and line activating the same code, and if that same code was used during the
subsequent one-to-one training, then presentation of a line sample during transfer testing
might tend to produce the same response as that produced by the color sample to which it
is functionally equivalent. Given the design specified in Figure 3, in transfer
testing, presentation of vertical would tend to produce the choice associated with a red
sample (i.e., circle), and presentation of horizontal would tend to produce the choice
associated with a green sample (i.e., dots). Such a tendency would produce positive
transfer (accuracy above 50%) in group consistent and negative transfer (accuracy below
50%) in group inconsistent. This is precisely what Urcuioli et al. (1989)
observed: During the first 16 trials of transfer testing, accuracy was 71.9% in
group consistent and 35.4% in group inconsistent. Grant and Spetch (1994) have
observed similar transfer effects in subjects initially trained in a many-to-one procedure
involving line and temporal (2- and 10-s light presentations) samples.
Although such transfer results
provide strong support for the notion that functionally equivalent samples in a
many-to-one mapping activate the same code, these results are not informative in regard to
the content of the code. Roitblat's (1980; Roitblat & Scopatz, 1983) research
employing standard matching procedures encourages the view that the code content in
many-to-one procedures, at least those employing colors and lines as stimuli, is
prospective. Nonetheless, as pointed out some time ago (Grant, 1982a), the data from
experiments employing colors and lines in many-to-one procedures can be explained
adequately by the notion that functionally equivalent samples activate the same code,
regardless of the content of that code. It is possible, for example, that samples
associated with a vertical choice stimulus activate a retrospective "sample A"
code and samples associated with a horizontal choice stimulus activate a retrospective
"sample B" code. It is also possible that samples associated with a
vertical choice stimulus active a prospective "vertical" (or "peck
vertical") code and samples associated with a horizontal choice stimulus activate a
prospective "horizontal" (or "peck horizontal") code.
Indeed, there is evidence that, at
least under some conditions involving "nonstandard" samples (such as the
occurrence and nonoccurrence of food), many-to-one training results in codes that do not
have prospective content (Kelly & Grant, 1998a; Urcuioli et al., 1994). In our
view, the results with nonstandard samples do not discourage the likely possibility that
many-to-one training with standard samples (i.e., colors, lines, and forms) results in
codes having prospective or anticipatory content. Nonetheless, it is our view that
studies employing many-to-one procedures, although often providing evidence consistent
with use of prospective codes, do not provide conclusive evidence for prospective code
content.
In
a spatial task. Particularly definitive evidence for
prospective coding was obtained by Zentall, Steirn, and Jackson-Smith (1990) who developed
a task to assess memory for spatial location in pigeons. An array of five pecking
keys was illuminated at the onset of a trial. Choosing (i.e., pecking) a particular
key was correct if that key had not been chosen previously on that trial. An error
consisted of choosing a key which had been previously chosen on that trial, and resulted
in a 2.5-s blackout after which the keys were again illuminated for a choice. A
trial ended when all five keys had been chosen.
The data of relevance to the issue
of prospective code content came from trials in which a delay was inserted at some point
in the choice sequence. Across trials, the delay occurred after 1, 2, 3, or 4
correct choices had been made. Zentall et al. (1990) found that the delay caused
more errors when it was interpolated after choice 2 than when it was interpolated after
choice 1. This result suggests that the amount of information that the bird had to
retain was greater after having made two choices than after having made only one
choice. Hence, this result suggests that the animal begins the trial using a code
having retrospective content; that is, early in the trial the animal remembers which
locations it has previously chosen. Interestingly, however, a delay interpolated
after choice 3 produced approximately the same amount of forgetting as a delay
interpolated after choice 2, suggesting that the memory load was equivalent after either
two or three choices had been made. Equivalent memory load after two or three
choices would obtain if pigeons remembered retrospectively after having made two choices
(and hence remembered the two locations previously chosen) and remembered prospectively
after having made three choices (and hence remembered the two locations not yet
chosen).
Further support for the notion that
pigeons switched to prospective coding later in the trial was provided by the finding that
a delay interpolated after choice 4 produced less forgetting than a delay interpolated
after choice 2 or 3 and, moreover, produced about the same amount of forgetting as a delay
interpolated after choice 1. This result is expected if the animal codes prospectively
later in the trial because the memory load after choice 4 (i.e., 1 item) is equivalent to
that after choice 1 and is less than that after choice 2 or 3 (i.e., 2 items).
Hence, the results of Zentall et al.'s (1990) experiment provides strong evidence that
codes having prospective or anticipatory content can mediate retention in pigeons (for a
conceptually similar experiment employing rats, see Cook, Brown, & Riley, 1985).
Summary
The data reviewed in this section
provide some support for the notion that codes having prospective or anticipatory content
can mediate short-term retention in pigeons. The strongest of such evidence is
provided by the research of Zentall et al. (1990) using a spatial task. Evidence
from other procedures is, at best, suggestive of prospective mediation. In
particular, although Roitblat's (1980) analysis of confusion errors in standard delayed
matching provides evidence for mediation of short-term retention by codes having
prospective or anticipatory content, that research involved a small sample size (i.e.,
three) and the effects suggesting prospective coding were significant only in two of the
three subjects. A replication of Roitblat's experiment would be welcome.
Moreover, the results of Roitblat and Scopatz's (1983) analysis of interference can, in
our view, be interpreted without maintaining that prospective coding was mediating
retention. Finally, data from many-to-one matching procedures involving colors,
lines, and/or forms support the view that functionally equivalent samples activate the
same code. Although these procedures may result in birds using codes having
prospective content, the data do not demand such an interpretation. In our view,
therefore, only the data from Zentall et al.'s experiment provide definitive evidence for
mediation of short-term retention in pigeons by codes having anticipatory content.
In the next two
sections we consider whether other types of anticipations can mediate short-term
retention. Specifically, we consider whether anticipated trial outcomes (Section IV)
and anticipated response patterns (Section V) can mediate short-term retention in
pigeons.
IV. Outcome
Expectancies and Short-Term Retention
The Differential
Outcome Procedure
In the matching procedures
considered so far in this chapter, correct responses are always reinforced by the same
outcome, typically 3-s access to grain. In the differential outcome procedure, a
procedure developed by Trapold (1970), this is not the case. Peterson and his
associates adapted a differential outcome procedure to the delayed matching task with
pigeons (Brodigan & Peterson, 1976; Peterson, Wheeler, & Armstrong, 1978).
In their procedure, color sample stimuli were followed by line choice stimuli and,
importantly, correct responses produced one of two outcomes, food or water, with equal
probability. As illustrated in Figure
4,  in the differential outcome group, the type of
outcome was perfectly correlated with type of sample and correct choice stimulus.
Hence, for example, choice of vertical after a red sample always produced grain, whereas
choice of horizontal after a green sample always produced water. In the
nondifferential outcome group, there was no correlation between sample/choice stimuli and
outcome. That is, choice of vertical after a red sample produced food and water
equally often, and, similarly, choice of horizontal after a green sample produced food and
water equally often. in the differential outcome group, the type of
outcome was perfectly correlated with type of sample and correct choice stimulus.
Hence, for example, choice of vertical after a red sample always produced grain, whereas
choice of horizontal after a green sample always produced water. In the
nondifferential outcome group, there was no correlation between sample/choice stimuli and
outcome. That is, choice of vertical after a red sample produced food and water
equally often, and, similarly, choice of horizontal after a green sample produced food and
water equally often.
The research discussed in Section I of
this article suggests that pigeons learn to anticipate trial outcomes after identifying
the sample. Hence, birds in the differential outcome group would learn to expect
food if the sample is identified as red (because correct choice on red-sample trials
produces food) and to expect water if the sample is identified as green (because correct
choice on green-sample trials produces water). Notice that these pigeons have two
potential sources of control over choice responding: a code (either retrospective or
prospective) and an outcome expectancy. With regard to the latter, an expectancy for
food should lead to choice of vertical because choice of vertical is always (and choice of
horizontal is never) reinforced in the presence of a food expectancy. An expectancy
for water, on the other hand, should lead to choice of horizontal because choice of
horizontal is always (and choice of vertical is never) reinforced in the presence of a
water expectancy.
Notice that in the nondifferential
outcome group, this second source of control over choice responding is not available
because the same expectancy should be present on all trials. Pigeons in this group
should expect both food and water regardless of whether the sample is identified as red or
green because both types of trials terminate, after a correct choice, in food and water
equally often. Hence, the same expectancy is present on trials in which choice of
vertical is reinforced and on trials in which choice of horizontal is reinforced.
Therefore, the expectancy provides no basis for responding differentially to the choice
stimuli.
Numerous pairs of outcomes in
addition to food and water have been employed in differential outcome experiments.
These pairs have included different types of food (e.g., Edwards, Jagielo, Zentall, &
Hogan, 1982), food and tone (e.g., Peterson, Wheeler, & Trapold, 1980), different
nonzero probabilities of food (e.g., Santi & Roberts, 1985), and illuminated
presentations of food and no food (e.g., Peterson et al., 1980; Urcuioli, 1990; Zentall
& Sherburne, 1994). Regardless of which type of outcome pair is employed,
matching performance in a differential outcome group is typically superior to that in a
nondifferential outcome group. This finding, known as the differential outcome
effect, is illustrated, and its theoretical significance considered, in the next
section.
The Differential Outcome Effect
As noted in the previous section, use
of differential outcomes typically enhances matching performance. This enhancement
is manifest in two ways. Specifically, use of differential outcomes facilitates
acquisition of the matching task and also enhances retention when delays are interpolated
between sample termination and presentation of choice stimuli. In general, the term
"differential outcome effect" is used to refer to either or both of these
aspects of facilitation of matching performance.
A recent experiment conducted in our
laboratory provides a demonstration of the differential outcome effect. Kelly and
Grant (1998b) trained pigeons on an identity-matching task using horizontal and vertical
stimuli. A 0-s delay intervened between sample termination and onset of the choice
stimuli (i.e., choice stimuli were presented immediately after termination of the sample).
Correct responses were followed by 3-s of illumination of the feeder, accompanied either
by access to food (hereafter referred to simply as "food") or no access to food
(hereafter referred  to simply
as "no food"). Incorrect responses were followed by nothing (i.e., the
feeder was not illuminated and food was not available). As illustrated in
Figure 5, sample type (and hence
correct choice) was correlated with outcome in the differential outcome group. In
the nondifferential outcome group, sample type (and hence correct choice) and outcome were
not correlated. to simply
as "no food"). Incorrect responses were followed by nothing (i.e., the
feeder was not illuminated and food was not available). As illustrated in
Figure 5, sample type (and hence
correct choice) was correlated with outcome in the differential outcome group. In
the nondifferential outcome group, sample type (and hence correct choice) and outcome were
not correlated.
Figure 6 shows matching accuracy in group
differential and nondifferential during the first 16 sessions of training. A
differential outcome 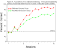 effect is apparent in that accuracy was higher in the
differential than in the nondifferential outcome group. Sessions to meet an
acquisition criterion of two consecutive sessions in which accuracy was at least 80%
correct with both types of samples also suggested an enhancement in acquisition in the
differential outcome group. Specifically, the nondifferential outcome group required
a mean of 24.6 sessions to met the acquisition criterion, whereas the differential outcome
group required a mean of only 15.4 sessions. effect is apparent in that accuracy was higher in the
differential than in the nondifferential outcome group. Sessions to meet an
acquisition criterion of two consecutive sessions in which accuracy was at least 80%
correct with both types of samples also suggested an enhancement in acquisition in the
differential outcome group. Specifically, the nondifferential outcome group required
a mean of 24.6 sessions to met the acquisition criterion, whereas the differential outcome
group required a mean of only 15.4 sessions.
After all animals were responding
accurately at a 0-s delay, a retention test was conducted in which delays of 0, 2, or 6 s
intervened between sample termination and onset of the choice stimuli. The data from
this test, shown in Figure 7, revealed
the second aspect of the differential outcome effect: enhanced retention in animals
receiving differential outcomes. between sample termination and onset of the choice stimuli. The data from
this test, shown in Figure 7, revealed
the second aspect of the differential outcome effect: enhanced retention in animals
receiving differential outcomes.
The occurrence of the differential
outcome effect and, especially, the facilitation of retention by differential outcomes,
suggests that short-term retention performance can be mediated by an outcome
expectancy. Moreover, as the data in Figure 7 reveal, these outcome expectancies
appear to be particularly memorable, and hence are available to guide choice responding
after the code activated by the sample is no longer active. In the next section, we
consider whether outcome expectancies need necessarily involve affectively potent events
in order to enhance acquisition and retention in delayed matching.
Differential Color Outcomes
Kelly and Grant (1998b) performed an
experiment similar to that described in the preceding section. As in the previous
experiment, pigeons were trained on an identity matching task using horizontal and
vertical stimuli, and correct responses were followed equally often by one of two
outcomes. However, the outcomes employed were a 1.5-s presentation of blue or yellow
light.  Following a correct choice, the choice stimuli were terminated and a color
was projected onto the key that the bird had chosen. Termination of the color was
followed by 3-s access to food. As
illustrated in Figure 8, sample type
(and hence correct choice) was correlated with outcome in the differential outcome
group. In the nondifferential outcome group, sample type (and hence correct choice)
and outcome were not correlated. Following a correct choice, the choice stimuli were terminated and a color
was projected onto the key that the bird had chosen. Termination of the color was
followed by 3-s access to food. As
illustrated in Figure 8, sample type
(and hence correct choice) was correlated with outcome in the differential outcome
group. In the nondifferential outcome group, sample type (and hence correct choice)
and outcome were not correlated.
Figure 9 shows matching accuracy in group
differential and nondifferential during the first 16 sessions of training. As was
the case when 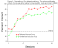 the outcomes were food and no food, a differential outcome
effect is apparent in that accuracy was higher in the differential than in the
nondifferential outcome group. Sessions to meet an acquisition criterion of two
consecutive sessions in which accuracy was at least 80% correct with both types of samples
also suggested an enhancement in acquisition in the differential outcome group.
Specifically, the nondifferential outcome group required a mean of 13.0 sessions to met
the acquisition criterion, whereas the differential outcome group required a mean of only
8.3 sessions. the outcomes were food and no food, a differential outcome
effect is apparent in that accuracy was higher in the differential than in the
nondifferential outcome group. Sessions to meet an acquisition criterion of two
consecutive sessions in which accuracy was at least 80% correct with both types of samples
also suggested an enhancement in acquisition in the differential outcome group.
Specifically, the nondifferential outcome group required a mean of 13.0 sessions to met
the acquisition criterion, whereas the differential outcome group required a mean of only
8.3 sessions.
Figure
10 shows the results of two retention tests. The first test, using delays of 0,
.3, and 1 s,  was conducted immediately following acquisition of 0-s delayed
matching. The second test, employing delays of 1, 5, and 9 s was conducted after
extensive training with a fixed delay of 1 s on all trials. In neither case was
there evidence that differential outcomes enhanced retention. In fact, there was a
slight tendency in the opposite direction. was conducted immediately following acquisition of 0-s delayed
matching. The second test, employing delays of 1, 5, and 9 s was conducted after
extensive training with a fixed delay of 1 s on all trials. In neither case was
there evidence that differential outcomes enhanced retention. In fact, there was a
slight tendency in the opposite direction.
In an effort to increase the
salience of the color outcomes we increased their duration from 1.5 s to 5.0 s, and
reduced the probability of food following correct choices from 1.0 to 0.5. Moreover,
retention tests were conducted after training at various nonzero fixed delays. In
spite of these manipulations, we have generally observed overlapping retention functions
in the two groups. To date, therefore, we have obtained no evidence that
differential color outcomes enhance short-term retention.
Summary
Research using differential outcome
procedures has provided evidence that pigeons learn to anticipate affective
outcomes. If the outcomes are differential with regard to the combination of sample
and correct choice, then sample presentation triggers an expectation of the outcome
associated with that sample. Data from retention testing reveal that these
expectations are particularly effective in mediating short-term retention. To date,
however, we have not obtained evidence that pigeons learn to anticipate more affectively
neutral outcomes (i.e., different colors). The finding that differential color
outcomes enhance acquisition could be explained without postulating that birds in the
differential outcome group learn to anticipate the color outcome. Specifically,
differential color outcomes may enhance the discriminability of the line sample stimuli, a
phenomenon known as acquired distinctiveness of cues. Early in training, at
which point the ability to distinguish between the lines is still developing, enhancing
the discriminability of the lines by differentially associating them with color outcomes
would facilitate performance and increase rate of acquisition. After extensive
training, the ability to discriminate between the lines based on their visual properties
may be sufficient to allow accurate identification of the sample on all trials. At
this point, enhancing the discriminability of the lines by associating them with different
colors would no longer enhance the likelihood of correct sample identification.
Although our data do not provide
evidence that pigeons can anticipate color outcomes, they also do not, of course, rule out
that possibility. In particular, it is possible that birds in the differential
outcome group did learn to anticipate the color outcomes, but that those anticipations
failed to facilitate retention. It may be that the anticipation of a color, in
contrast to the anticipation of an affective event, is not more memorable than the code
activated by the sample. If so, no enhancement in retention would occur in spite of
the fact that birds in a differential outcome group learn to anticipate the color
outcome. Research is continuing in our laboratory to determine whether there are
conditions that generate expectancies for more affectively neutral events that function as
effective mediators of short-term retention.
V. "Simple" Response Intentions and
Short-Term Retention
Simple vs.
Conditional Discrimination Procedures
As discussed previously in this
article, matching-to-sample is a conditional discrimination task because the reinforcement
contingencies vary from trial to trial. In a simple discrimination task, in
contrast, the reinforcement contingencies remain constant from trial to trial. For
example, pecking a horizontal line might always produce reinforcement whereas pecking a
vertical line never produces reinforcement.
Honig and Wasserman (1981) modified
the delayed matching procedure and a simple discrimination procedure in order to compare
retention in the two tasks. As developed in more detail below, performance in a
delayed simple discrimination can be mediated by a "simple" response intention;
that is, to peck or not peck the test stimulus. Hence, if pigeons are capable of
anticipating a future action, and if such an anticipation is an effective mediator of
short-term retention, then retention should be higher in a delayed simple discrimination
than in a delayed conditional discrimination.
In Honig and Wasserman's (1981)
procedure, trials began with presentation of an initial stimulus that, after a delay
interval, was followed by presentation of a single test stimulus. On some trials the
test stimulus was positive and the first peck after 5 s in the presence of the test
stimulus produced reinforcement (i.e., fixed interval, FI, 5 s schedule). On other
trials the test stimulus was negative and terminated in nonreinforcement after 5 s (i.e.,
fixed time, FT, 5 s extinction schedule).
As pigeons learn to discriminate
positive and negative test stimuli, rate of keypecking becomes increasingly
differential: high in the presence of positive test stimuli and low in the presence
of negative test stimuli. The extent of differential responding is indexed by a
discrimination ratio that represents the proportion of responses to all test stimuli that
are emitted to positive test stimuli. Thus, a ratio of .5 indicates equivalent
responding to, and hence no discrimination between, positive and negative test
stimuli. Ratios between .5 and 1.0 indicate increasingly higher levels of
discrimination.
In Honig and Wasserman's (1981)
experiment, the simple and conditional procedures differed in  terms of the basis for
determining the positive or negative status of a test stimulus. In the delayed
simple discrimination,
illustrated in the left half of Figure 11,
the initial stimulus provided all the information necessary to determine whether pecking
the test
stimulus would or would not be reinforced. In the case illustrated, a
red initial stimulus signalled that the test stimulus was positive, regardless of whether
that test stimulus was a vertical or horizontal line. A blue initial stimulus, in
contrast, signalled that the test stimulus was negative, regardless of whether that test
stimulus was a vertical or horizontal line.
Click for an animated illustration of the
delayed simple discrimination procedure terms of the basis for
determining the positive or negative status of a test stimulus. In the delayed
simple discrimination,
illustrated in the left half of Figure 11,
the initial stimulus provided all the information necessary to determine whether pecking
the test
stimulus would or would not be reinforced. In the case illustrated, a
red initial stimulus signalled that the test stimulus was positive, regardless of whether
that test stimulus was a vertical or horizontal line. A blue initial stimulus, in
contrast, signalled that the test stimulus was negative, regardless of whether that test
stimulus was a vertical or horizontal line.
Click for an animated illustration of the
delayed simple discrimination procedure
In the delayed conditional
discrimination, illustrated in the right half of Figure 11, the combination of
initial and test stimulus determined whether pecking the test stimulus would or would
not
be reinforced. As illustrated, vertical was positive after violet but negative after
yellow, whereas horizontal was positive after yellow and negative after violet. Two sets of initial stimuli were employed to allow a
within-subjects comparison of retention in the two tasks (i.e., all subjects were trained
and tested on both tasks). Click
for an animated illustration of the delayed conditional discrimination procedure
Notice that because the identity of the
test stimulus is irrelevant in the simple discrimination task, retention could be mediated
by a "simple" response intention. That is, birds could remember to peck on
trials initiated by red, and not to peck on trials initiated by blue. However,
because the identity of the test stimulus is relevant in the conditional discrimination
task, mediation by a simple response intention is precluded. Hence, if pigeons can
anticipate a future action (i.e., pecking or not pecking), then one might expect better
retention in the simple than in the conditional task. This prediction is based on
the notion that a simple response intention (i.e., "peck" or "do not
peck") is more memorable than is either a prospective code (e.g., "peck
vertical" or "peck horizontal") or a retrospective code (e.g.,
"violet" and "yellow"). Tests of this prediction are considered
in the next section.
Retention in Simple and Conditional Procedures
 Following acquisition, Honig and
Wasserman (1981) assessed retention in the two tasks. As shown in Figure 12, retention in the simple task
was considerably more robust than that in
the conditional task (the data points in Figure 12 were estimated from Figure 4 in Honig
& Wasserman, 1981). This finding is consistent with the notion that in the
simple task, pigeons remembered an anticipated action (to peck or not peck the test
stimulus). Following acquisition, Honig and
Wasserman (1981) assessed retention in the two tasks. As shown in Figure 12, retention in the simple task
was considerably more robust than that in
the conditional task (the data points in Figure 12 were estimated from Figure 4 in Honig
& Wasserman, 1981). This finding is consistent with the notion that in the
simple task, pigeons remembered an anticipated action (to peck or not peck the test
stimulus).
Although Honig and Wasserman's
(1981) findings are consistent with the view that simple response intentions mediate
performance in a delayed simple discrimination, their findings do not demand such an
account. As they noted, enhanced retention in the simple task could be an instance
of the differential outcome effect. Notice that in the simple task, the initial
stimuli are correlated with different outcomes. In the illustration shown in
Figure 11, red is associated with a food
outcome and blue with a no-food outcome. Hence, red should come to evoke an
expectation of food and blue should come to evoke an expectation of no food. It may
be that these expectations persisted throughout the retention interval and controlled
responding to the test stimuli. Specifically, an expectation for food evoked pecking
of the test stimulus, and an expectation for no food failed to evoke pecking. In the
conditional task, the initial stimuli are not correlated with different outcomes and hence
should evoke the same expectancy (i.e., an expectancy for food). As discussed in
Section IVb, anticipated affective outcomes are particularly effective mediators of
short-term retention in pigeons. Hence, enhanced retention in the simple task could
reflect mediation of retention in that task by differential outcome expectancies.
Urcuioli and Zentall (1990) devised
a clever design which allowed them to assess whether the enhanced retention in the simple
task obtained by Honig and Wasserman (1981) reflected mediation by simple response
intentions or mediation by outcome expectancies. They modified the simple task such
that all trials, regardless of which initial stimulus was presented, ended in
reinforcement. The effect of this modification, of course, is to remove the
correlation between initial stimulus and  outcome. As
shown in the left half of Figure 13,
the initial stimuli in the simple task signalled not whether the trial would end in food
or no food, but rather which behavior, pecking or not pecking the test stimulus, was
required to procure reinforcement. Following a red initial stimulus, the first key
peck after 5 s in the presence of the test stimulus produced reinforcement (i.e., FI 5 s
schedule), regardless of whether the test stimulus was vertical or horizontal.
Following a green initial stimulus, in contrast, the bird had to refrain from pecking for
5 s to produce reinforcement (i.e., differential reinforcement of other behavior, DRO, 5 s
schedule). As in the simple task employed by Honig and Wasserman (1981), the initial
stimuli in Urcuioli and Zentall's symmetrically-reinforced simple task supplied all the
information necessary to determine the behavior required at testing. Importantly
however, and in contrast to Honig and Wasserman's simple task, the initial stimuli were
not correlated with differential outcomes in Urcuioli and Zentall's simple
task. Hence, retention in this task could be
mediated by simple response intentions (i.e., peck, do not peck), but could not be
mediated by differential outcome expectancies. Click for an animated illustration of a
symmetrically-reinforced delayed simple discrimination outcome. As
shown in the left half of Figure 13,
the initial stimuli in the simple task signalled not whether the trial would end in food
or no food, but rather which behavior, pecking or not pecking the test stimulus, was
required to procure reinforcement. Following a red initial stimulus, the first key
peck after 5 s in the presence of the test stimulus produced reinforcement (i.e., FI 5 s
schedule), regardless of whether the test stimulus was vertical or horizontal.
Following a green initial stimulus, in contrast, the bird had to refrain from pecking for
5 s to produce reinforcement (i.e., differential reinforcement of other behavior, DRO, 5 s
schedule). As in the simple task employed by Honig and Wasserman (1981), the initial
stimuli in Urcuioli and Zentall's symmetrically-reinforced simple task supplied all the
information necessary to determine the behavior required at testing. Importantly
however, and in contrast to Honig and Wasserman's simple task, the initial stimuli were
not correlated with differential outcomes in Urcuioli and Zentall's simple
task. Hence, retention in this task could be
mediated by simple response intentions (i.e., peck, do not peck), but could not be
mediated by differential outcome expectancies. Click for an animated illustration of a
symmetrically-reinforced delayed simple discrimination
To maintain comparability between the
simple and conditional tasks, the conditional task was modified so that all trials ended
in reinforcement, as was the case in the simple task. Also as in the simple task,
the response required to produce reinforcement varied from trial-to-trial (see right half
of Figure 13). Notice, however,
that in the symmetrically-reinforced conditional task it was the combination of initial and test stimulus that determined whether pecking or not
pecking was reinforced on the current trial. Hence, performance in the conditional
task could not be mediated by a simple response intention. Click for an animated illustration of a
symmetrically-reinforced delayed conditional discrimination
Thus, if simple response intentions
mediate retention in the simple task, then Urcuioli and Zentall's (1990) procedure should
reveal enhanced retention in the simple task, as did Honig and Wasserman's (1981)
procedure. If, however, the facilitation in retention observed in the simple task by
Honig and Wasserman was mediated exclusively by differential outcome expectancies, then
Urcuioli and Zentall's procedure should fail to reveal enhanced retention in the simple
task.
Following acquisition, two retention
tests were conducted. The first involved retention intervals of 0, 5, and 10 s, and
the second  involved retention intervals of 0, 10, and 20 s. The results of the retention tests are
shown in Figure 14 (the data points
plotted in Figure 14 were estimated from Figure 3 in Urcuioli & Zentall, 1990).
Although overall accuracy was somewhat higher in the simple than in the conditional task,
this difference was not statistically significant in either retention test. More
importantly, and in contrast to the findings of Honig and Wasserman (1981), there was no
evidence of enhanced retention in the simple task. In fact, rate of forgetting was
slightly faster in the simple than conditional task, although this difference was not
statistically significant.
involved retention intervals of 0, 10, and 20 s. The results of the retention tests are
shown in Figure 14 (the data points
plotted in Figure 14 were estimated from Figure 3 in Urcuioli & Zentall, 1990).
Although overall accuracy was somewhat higher in the simple than in the conditional task,
this difference was not statistically significant in either retention test. More
importantly, and in contrast to the findings of Honig and Wasserman (1981), there was no
evidence of enhanced retention in the simple task. In fact, rate of forgetting was
slightly faster in the simple than conditional task, although this difference was not
statistically significant.
Urcuioli and Zentall's (1990)
results suggest that the enhanced retention in the simple task, compared to that in the
conditional task, obtained by Honig and Wasserman (1981) was due entirely to mediation of
retention by differential outcome expectancies in the simple task. However, as
Urcuioli and Zentall (1992) noted, the results of their 1990 study (illustrated in
Figure 14) do not rule out the possibility
that response intentions did mediate performance in the delayed simple
discrimination. To account for equivalent rates of forgetting in simple and
conditional tasks, one would be required to assume that a simple response intention is no
more (and no less) memorable than the code that mediates retention in a delayed
conditional discrimination (i.e., a retrospective or prospective code).
To further pursue the possibility
that response intentions mediate performance in delayed simple discriminations, Urcuioli
and Zentall (1992) employed transfer tests. In one test, illustrated in
Figure 15, pigeons were trained
sequentially on two simple  discriminations involving different initial and test
stimuli. Following acquisition, the initial stimuli from the first task were
followed by the test stimuli from the second task. If performance is mediated by
simple response intentions, then considerable transfer would be expected.
Specifically, if red evokes the anticipation "peck", then a high rate of pecking
the blue and white test stimuli should occur on transfer trials initiated by red.
Further, if green evokes the anticipation "do not peck", then a low rate of
pecking the blue and white test stimuli should occur on transfer trials initiated by
green. In contrast to this prediction, red and green failed to control different
rates of pecking the blue and white test stimuli. Thus, results of transfer testing
also failed to provide evidence of mediation of short-term retention by response
intentions. discriminations involving different initial and test
stimuli. Following acquisition, the initial stimuli from the first task were
followed by the test stimuli from the second task. If performance is mediated by
simple response intentions, then considerable transfer would be expected.
Specifically, if red evokes the anticipation "peck", then a high rate of pecking
the blue and white test stimuli should occur on transfer trials initiated by red.
Further, if green evokes the anticipation "do not peck", then a low rate of
pecking the blue and white test stimuli should occur on transfer trials initiated by
green. In contrast to this prediction, red and green failed to control different
rates of pecking the blue and white test stimuli. Thus, results of transfer testing
also failed to provide evidence of mediation of short-term retention by response
intentions.
The failure of Urcuioli and Zentall
(1990, 1992) to obtain evidence of mediation by simple response intentions does not, of
course, rule out the possibility that under some conditions such mediation might be
observed. Research in our laboratory has recently addressed this possibility.
As noted previously, the identity of
the test stimulus is irrelevant in a simple discrimination. Hence, only a single
test stimulus could be used and, for example, trials might involve reinforced pecking of a
white test stimulus after a red initial stimulus and reinforced nonpecking of white light
after green. Presumably, Honig and Wasserman (1981) and Urcuioli and Zentall (1990,
1992) used two test stimuli in the simple task to maintain comparability with the
conditional task which requires use of at least two different test stimuli. Notice,
however, that the possibility of mediation of retention by simple response intentions is
predicated on pigeons being sensitive to the irrelevance of test stimulus identity.
If, however, pigeons are insensitive to the irrelevance of test stimulus identity in a
simple task using two test stimuli, then mediation by simple response intentions would not
be expected.
Recent research in our laboratory
has addressed the possibility that pigeons fail to detect the irrelevance of test stimulus
identity when two test stimuli are employed in the simple task (Grant & Kelly, 1998;
Grant, Kelly, & Steinbring, 1997). In particular, we reasoned that the
irrelevance of test stimulus identity might be more readily detected if either (a) only a
single test stimulus was employed or (b)  many test stimuli were employed. Therefore,
we implemented a modification of the design employed by Urcuioli and Zentall (1990).
As illustrated in Figure 16, in
the two simple discrimination
groups, the initial stimulus supplied all the information necessary to determine which
behavior would be reinforced at the time of testing. In the one test stimulus group,
the test stimulus was always horizontal. In the many test stimuli group, the test
stimulus was equally often one of nine possible stimuli. Finally, the delayed
conditional group received training equivalent to that received by Urcuioli and Zentall's
(1990) conditional discrimination group. Grant and Kelly (1998) replicated the
procedure shown in Figure 16
except that (a) horizontal replaced red and vertical replaced green as initial stimuli and
(b) red replaced horizontal and green replaced vertical as test stimuli. many test stimuli were employed. Therefore,
we implemented a modification of the design employed by Urcuioli and Zentall (1990).
As illustrated in Figure 16, in
the two simple discrimination
groups, the initial stimulus supplied all the information necessary to determine which
behavior would be reinforced at the time of testing. In the one test stimulus group,
the test stimulus was always horizontal. In the many test stimuli group, the test
stimulus was equally often one of nine possible stimuli. Finally, the delayed
conditional group received training equivalent to that received by Urcuioli and Zentall's
(1990) conditional discrimination group. Grant and Kelly (1998) replicated the
procedure shown in Figure 16
except that (a) horizontal replaced red and vertical replaced green as initial stimuli and
(b) red replaced horizontal and green replaced vertical as test stimuli.
Following acquisition, birds received a
retention test involving delays of 0, 5 and 15 s. As shown in Figure 17, accuracy was slightly higher in
the simple task employing  one test stimulus than
in both the simple task employing nine test stimuli and the conditional task, although
this difference was not statistically significant. More importantly, neither
experiment produced evidence that rate of forgetting was reduced in either simple task
relative to that in the conditional task. Hence, neither experiment produced
evidence that simple response intentions can mediate short-term retention in pigeons. one test stimulus than
in both the simple task employing nine test stimuli and the conditional task, although
this difference was not statistically significant. More importantly, neither
experiment produced evidence that rate of forgetting was reduced in either simple task
relative to that in the conditional task. Hence, neither experiment produced
evidence that simple response intentions can mediate short-term retention in pigeons.
Summary
Recent research has
shown that an early report of enhanced retention in a delayed simple discrimination,
relative to that in a delayed conditional discrimination, was an instance of the
differential outcome effect. When the operation of a differential outcome effect in
the simple task is precluded, rate of forgetting in simple and conditional tasks is
equivalent. Hence, research has not revealed convincing evidence that short-term
retention in pigeons can be mediated by simple response intentions (i.e., peck, do not
peck).
VI. Conclusion
The question of interest in this
chapter is whether an anticipation of a future action and/or event can function as an
effective mediator of short-term retention in pigeons. Although the research
reviewed above permits that question to be answered in the affirmative, mediation of
retention by anticipated events may occur only in a limited set of circumstances.
Perhaps the strongest evidence for anticipatory memory-mediation comes from Zentall et
al.'s (1990) study investigating memory for spatial locations. Recall that they
found that a delay interpolated after four of five locations had been chosen produced less
disruption in performance than a delay interpolated after two or three locations had been
chosen. This finding provides compelling evidence that, as the trial progressed,
pigeons recoded from remembering locations chosen to remembering locations not yet chosen.
Research using differential outcomes
also appears to provide strong evidence for anticipatory memory-mediation.
Specifically, retention is enhanced when correct responses following each sample result in
different outcomes. This finding suggests that the samples in such differential
outcome procedures evoke different outcome expectancies that persist during the delay and
control choice performance. However, one might ask about the nature of these
expectancies and, in particular, whether they are truly anticipatory. To elaborate,
consider the possibility that the expectancies take the form of a visual image of food
(e.g., an illuminated feeder with grain accessible) and a visual image of no food (e.g.,
darkness or illuminated feeder with grain inaccessible). Likely most readers would
agree that expectancies of this sort are truly anticipatory. However, consider that
the occurrence of food and no food elicit radically different emotional reactions in a
hungry pigeon. Therefore, for example, a red sample should evoke a positive
emotional reaction if correct responses on red-sample trials always result in food.
In contrast, a green sample should evoke a negative emotional reaction if correct
responses on green-sample trials always result in no food. Perhaps it is these
emotional reactions, unaccompanied by a representation of the outcome, which persist
during a delay and guide choice behavior. If so, is this a genuine instance of
anticipatory memory? In our view, it is not.
It may be argued that an account of
the differential outcome effect in terms of persisting emotions is flawed because the
different outcomes employed have not always involved one favorable and one unfavorable
outcome. For example, enhanced retention as a function of differential outcomes has
been observed when the outcomes have been different probabilities of food or different
types of appetitive reinforcers (e.g., different types of food). Even in these
instances, however, the different outcomes are likely to evoke, at the least, different
magnitudes of the same affective state. In our view, it is reasonable that a bird
could learn to choose between two stimuli based upon its current level of
"excitement".
Research in our laboratory using
different colors as outcomes is consistent with an interpretation of the differential
outcome effect in terms of persisting emotional reactions. Because the color
outcomes are followed equally often by food, they should not elicit different types or
amounts of affect. Hence, samples associated differentially with those colors should
also not evoke emotional states which are different in either type or magnitude.
Hence, if facilitation of retention in a differential outcome procedure requires that the
samples evoke qualitatively or quantitatively different emotional reactions, then such
facilitation should not occur when those outcomes are colors with an equivalent historical
relation to food. Consistent with this supposition, we have not observed enhanced
retention by differential outcomes when those outcomes have been colored lights. It
should be noted, however, that other explanations of the failure for differential color
outcomes to enhance retention are possible. For example, pigeons may be capable of
anticipating outcomes only if those outcomes have strong affective properties or,
alternatively, perhaps pigeons do learn to anticipate color outcomes, but those
anticipations are no more memorable than retrospective or prospective codes.
Also reviewed in this article were
data from retention and transfer testing that failed to provide evidence that simple
response intentions mediate short-term retention in pigeons. Hence, pigeons appear
incapable of using an anticipated action (i.e., peck or do not peck) to mediate short-term
retention. Moreover, data from these experiments may have implications for the view
that standard matching tasks typically involve prospective mediation. Recall that
retention in our simple discrimination involving many test stimuli was equivalent to that
in the conditional discrimination. This finding suggests that pigeons in the former
group were not coding prospectively: that is, they were not remembering the nine test
stimuli to which responding should or should not be directed on each trial. Had this
been the case, retention would be expected to be markedly lower in the simple
discrimination with nine test stimuli than in the conditional discrimination because the
latter required only a single prospective code on each trial. One could argue, of
course, that birds in the many test stimuli group coded retrospectively and remembered
which color initiated each trial. Although this was likely the case, one might then
ask why retention was equivalent in this group and in the conditional discrimination
group, which, on the basis of Roitblat's (1980; Roitblat & Scopatz, 1983) research,
would be expected to code prospectively. Although one could argue that retrospective
codes of color stimuli and prospective codes of line stimuli are equally memorable, such a
notion strains credibility.
In our view, the most parsimonious
explanation of research comparing retention in simple and conditional discriminations is
that pigeons in all groups remembered the initial stimulus during the delay and then made
a decision about responding at the time of testing based on that retrospective code.
But if this explanation is correct, then how does one account for Roitblat's (1980;
Roitblat & Scopatz, 1983) data implicating prospective coding in choice
matching? One possibility is that conditional discriminations involving colors,
lines and forms are mediated by prospective codes when choice responding is involved, but
are mediated by retrospective codes when go/no go responding is involved (but see Grant
& Spetch, 1991; Spetch & Grant, 1993; Spetch, Grant, & Kelly, 1996).
Alternatively, perhaps prospective
coding was induced by presentation of three choice stimuli on each trial, which was the
case in both Roitblat (1980) and Roitblat and Scopatz (1983). It may be that the
utility of prospective coding, relative to that of retrospective coding, increases as the
number of choice stimuli that need to be identified and evaluated on each trial
increases. To illustrate, consider a situation involving five samples and five
choice stimuli, each of the latter being presented simultaneously at the time of
choice. If the animal codes retrospectively, it may be necessary to repeatedly
retrieve the response rule(s) associated with that code as each choice stimulus is
successively evaluated. The response decision process may be simplified if the
animal remembers prospectively, and hence need only determine whether a particular choice
stimulus matches the active code. Thus, there is some basis for suggesting that the
occurrence of prospective coding may be restricted to tasks, such as that employed by
Roitblat (1980) and Roitblat and Scopatz (1983), in which color or line samples are
followed by the simultaneous presentation of more than two choice stimuli.
In summary, the
literature provides only moderate evidence of a role for anticipation in mediation of
short-term retention in pigeons. The strongest evidence comes from a spatial task in
which pigeons appear able to remember locations not yet chosen. The differential
outcome literature is consistent with mediation by anticipated affective outcomes, but may
reflect instead persisting emotional or behavioral reactions elicited by samples
correlated with outcomes that differ affectively. Research using many-to-one
sample-to-choice mapping procedures is similarly not definitive with regard to whether the
codes mediating retention have anticipatory content. Moreover, research has failed
to obtain evidence that short-term retention can be mediated by either (a) anticipated
outcomes which are affectively equivalent or (b) simple response intentions.
Finally, although analysis of confusion errors and interference suggests prospective
coding in choice matching with lines or colors (at least in procedures involving the
simultaneous presentation of three choice stimuli), the small sample size and mixed
statistical evidence suggests caution in evaluating the theoretical significance of those
results. Continued research is necessary to clarify the role of anticipation as a
mediator of short-term retention in pigeons.
VII.
References
Brodigan, D. L., & Peterson, G.
B. (1976). Two-choice conditional discrimination
performance of pigeons as a function of reward expectancy. Animal Learning
& Behavior, 4, 121-124.
Colwill, R. M., & Rescorla, R.
A. (1985). Postconditioning devaluation of a reinforcer affects instrumental
responding. Journal of Experimental Psychology: Animal Behavior Processes,
11, 120-132.
Conrad, R. (1964). Acoustic
confusions in immediate memory. British Journal of Psychology, 55,
75-83.
Cook, R. G., Brown, M. F., &
Riley, D. A. (1985). Flexible memory processing by
rats: Use of prospective and retrospective information in the radial maze.
Journal of Experimental Psychology: Animal Behavior Processes, 11, 453-469.
Dickinson, A., Campos, J., Varga, Z.
L., & Balleine, B. (1996). Bidirectional instrumental
conditioning. Quarterly Journal of Experimental Psychology, 48B, 289-306.
Edwards, C. A., Jagielo, J. A.,
Zentall, T. R., & Hogan, D. E. (1982). Acquired eqivalence and distinctiveness
in matching to sample by pigeons: Mediation by reinforcer-specific
expectancies. Journal of Experimental Psychology: Animal Behavior
Processes, 8, 244-259.
Grant, D. S. (1981).
Short-term memory in the pigeon. In N. E. Spear & R. R. Miller (Eds.), Information
processing in animals: Memory mechanisms (pp. 227-256). Hillsdale, NJ:
Erlbaum.
Grant, D. S. (1982a).
Prospective versus retrospective coding of samples of stimuli, responses, and reinforcers
in delayed matching with pigeons. Learning and Motivation, 13, 265-280.
Grant, D. S. (1982b). Samples
of stimuli, responses, and reinforcers: Effect of incongruent sample type, serial
position, and mode of presentation. Animal Learning & Behavior, 10, 7-14.
Grant, D. S. (1991).
Symmetrical and asymmetrical coding of food and no-food samples in delayed matching in
pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 17,
186-193.
Grant, D. S., & Kelly, R.
(1998). [Failure to find evidence that response intentions mediate retention in
delayed simple discriminations with pigeons.] Unpublished raw data.
Grant, D. S.,
Kelly, R., & Steinbring, K. M. (1997, June). The role of response intentions in
delayed simple discriminations in pigeons. Presented at the meeting of the Canadian
Society for Brain, Behaviour, and Cognitive Science, University of Manitoba.
Grant, D. S., & Spetch, M. L.
(1991). Pigeons' memory for event duration: Differences between choice and
successive matching tasks. Learning and Motivation, 22, 180-199.
Grant, D. S., & Spetch, M. L.
(1993). Analogical and nonanalogical coding of samples differing in duration in a
choice-matching task in pigeons. Journal of Experimental Psychology: Animal
Behavior Processes, 19, 15-25.
Grant, D. S., & Spetch, M. L.
(1994). Mediated transfer testing provides evidence for common coding of duration
and line samples in many-to-one matching in pigeons. Animal Learning &
Behavior, 22, 84-89.
Holland, P. C., & Rescorla, R.
A. (1975). The effect of two ways of devaluing the
unconditioned stimulus after first- and second-order appetitive conditioning. Journal
of Experimental Psychology: Animal Behavior Processes, 1, 355-363.
Holland, P. C., & Straub, J. J.
(1979). Differential effect of two ways of devaluing
the unconditioned stimulus after Pavlovian appetitive conditioning. Journal
of Experimental Psychology: Animal Behavior Processes, 5, 65-78.
Honig, W. K., & Thompson, R. K.
R. (1982). Retrospective and prospective processing in animal working memory.
In G. H. Bower (Ed.), The psychology of learning and motivation: Advances in research
and theory (Vol. 16, pp. 239-283). New York, NY: Academic Press.
Honig, W. K., & Wasserman, E. A.
(1981). Performance of pigeons on delayed simple and conditional discrimination
under equivalent training procedures. Learning and Motivation, 12, 149-170.
Hull, C. L. (1943). Principles of
behavior. New York, NY: Appleton-Century-Crofts.
Hull, C. L. (1952). A Behavior system. New Haven, CT: Yale University Press.
Kamin, L. J. (1968).
"Attention-like" processes in classical conditioning. In M. R. Jones (Ed.), Miami
symposium on the prediction of behavior: Aversive stimulation (pp. 9-31).
Miami, FL:
University of Miami Press.
Kamin, L. J. (1969). Predictability,
surprise, attention, and conditioning. In B. A. Campbell & R. M. Church (Eds.), Punishment
and aversive behavior (pp. 279-296). New York, NY: Appleton-Century-Crofts.
Kelly, R., &
Grant, D. S. (1998a). Coding of duration and hedonic samples in many-to-one
matching-to-sample with pigeons. Manuscript submitted for publication.
Kelly, R., &
Grant, D. S. (1998b). [The effect of differential hedonic and nonhedonic outcome
procedures in delayed matching in pigeons]. Unpublished raw data.
Peterson, G. B., Wheeler, R. L.,
& Armstrong, G. D. (1978). Expectancies as mediators in the differential-reward
conditional discrimination performance of pigeons. Animal Learning &
Behavior, 6, 279-283.
Peterson, G. B., Wheeler, R. L.,
& Trapold, M. A. (1980). Enhancement of pigeons' conditional discrimination
performance by expectancies of reinforcement and nonreinforcement. Animal
Learning & Behavior, 8, 22-30.
Rescorla, R. A. (1966).
Predictability and number of pairings in Pavlovian fear conditioning. Psychonomic
Science, 4, 383-384.
Rescorla, R. A. (1968). Probability
of shock in the presence and absence of CS in fear conditioning. Journal of Comparative
and Physiological Psychology, 66, 1-5.
Riley, D. A., Cook, R. G., &
Lamb, M. R. (1981). A classification and analysis of short-term retention
codes. In G. H. Bower (Ed.), The psychology of learning and motivation: Advances
in research and theory (Vol. 15, pp. 51-79). New York, NY: Academic Press.
Roitblat, H. L. (1980). Codes
and coding processes in pigeon short-term memory. Animal Learning & Behavior,
8, 341-351.
Roitblat, H. L. (1993).
Representations and processes in working memory. In T. R. Zentall (Ed.), Animal
cognition: A tribute to Donald A. Riley (pp. 175-192). Hillsdale, NJ: Erlbaum.
Roitblat, H. L., & Scopatz, R.
A. (1983). Sequential effects in pigeon delayed
matching-to-sample performance. Journal of Experimental Psychology:
Animal Behavior Processes, 9, 202-221.
Santi, A., & Roberts, W. A.
(1985). Prospective representation: The effects of varied mapping of sample
stimuli to comparison stimuli and differential trial outcomes on pigeons' working
memory. Animal Learning & Behavior, 13, 103-108.
Spear, N. E., & Riccio, D. C.
(1994). Memory: Phenomena and principles. Boston, MA: Allyn and Bacon.
Spence, K. W. (1956). Behavior
theory and conditioning. New Haven, CT: Yale University Press.
Spence, K. W. (1960). Behavior
theory and learning. Englewood Cliffs, NJ: Prentice-Hall.
Spetch, M. L., & Grant, D. S.
(1993). Pigeons' memory for event duration in choice and successive
matching-to-sample tasks. Learning and Motivation, 24, 156-174.
Spetch, M. L., Grant, D. S., &
Kelly, R. (1996). Procedural determinants of coding processes n pigeons' memory for
duration. Learning and Motivation, 27, 179-199.
Thorndike, E. L. (1898). Animal
intelligence: An experimental study of the associative processes in animals.
Psychological
Review Monographs, 2(8).
Tolman, E. C. (1932). Purposive
behavior in animals and men. New York, NY: Century.
Tolman, E. C. (1948).
Cognitive maps in rats and men. Psychological Review, 55, 189-208.
Tolman, E. C., & Honzik, C. H.
(1930a). "Insight" in rats. University of California Publications
in Psychology, 4, 215-232.
Tolman, E. C., & Honzik, C. H.
(1930b). Introduction and removal of reward and maze performance in rats. University
of California Publications in Psychology, 4, 257-275.
Tolman, E. C., Ritchie, B. F., &
Kalish, D. (1946a). Studies in spatial learning I. Orientation and the short-cut. Journal
of Experimental Psychology, 36, 13-24.
Tolman, E. C., Ritchie, B. F., &
Kalish, D. (1946b). Studies in spatial learning II. Place learning versus response
learning. Journal of Experimental Psychology, 36, 221-229.
Trapold, M. A. (1970). Are
expectancies based upon different positive reinforcing events discriminably
different? Learning and Motivation, 1, 129-140.
Urcuioli, P. J. (1990).
Differential outcomes and many-to-one matching: Effects of correlation with correct
choice. Animal Learning & Behavior, 18, 410-422.
Urcuioli, P. J., DeMarse, T., &
Zentall, T. R. (1994). Some properties of many-to-one matching with hue, response,
and food samples: Retention and mediated transfer. Learning and Motivation, 25,
175-200.
Urcuioli, P. J., & Zentall, T.
R. (1986). Retrospective coding in pigeons' delayed
matching-to-sample. Journal of Experimental Psychology: Animal Behavior
Processes, 12, 69-77.
Urcuioli, P. J., & Zentall, T.
R. (1990). On the role of trial outcomes in delayed
discriminations. Animal Learning & Behavior, 18, 141-150.
Urcuioli, P. J., & Zentall, T.
R. (1992). Transfer across delayed
discriminations: Evidence regarding the nature of prospective working memory.
Journal of Experimental Psychology: Animal Behavior Processes, 18, 154-173.
Urcuioli, P. J., Zentall, T. R.,
Jackson-Smith, P., & Steirn, J. N. (1989). Evidence for common coding in
many-to-one matching: Retention, intertrial interference, and transfer. Journal
of Experimental Psychology: Animal Behavior Processes, 15, 264-273.
Wagner, A. R. (1969). Stimulus
validity and stimulus selection in associative learning. In N. J. Mackintosh &
W. K. Honig (Eds.), Fundamental issues in associative learning (pp. 90-122).
Halifax, Nova Scotia: Dalhousie University Press.
Wagner, A. R., Logan, F. A.,
Haberlandt, K., & Price, T. (1968). Stimulus
selection in animal discrimination learning. Journal of Experimental
Psychology, 76, 171-180.
Zentall, T. R., Jagielo, J. A.,
Jackson-Smith, P., & Urcuioli, P. J. (1987). Memory codes in pigeon short-term
memory: Effects of varying the number of samples and comparison stimuli. Learning
and Motivation, 18, 21-33.
Zentall, T. R., & Sherburne, L.
M. (1994). Role of differential sample responding in
the differential outcomes effect involving delayed matching by pigeons.
Journal
of Experimental Psychology: Animal Behavior Processes, 20, 390-401.
Zentall, T. R., Steirn, J. N., &
Jackson-Smith, P. (1990). Memory strategies in pigeons' performance of a
radial-arm-maze analog task. Journal of Experimental Psychology: Animal
Behavior Processes, 16, 358-371.
Zentall, T. R., Urcuioli, P. J., Jagielo, J. A., & Jackson-Smith, P. (1989). Interaction of sample dimension and
sample-comparison mapping on pigeons' performance of delayed conditional
discriminations. Animal Learning & Behavior, 17, 172-178.
|
Acknowledgement
Preparation of this chapter was
supported by a grant from the Natural Sciences and Engineering Research Council of Canada
awarded to the first author. Correspondence concerning this article should be sent
to: Douglas S. Grant, Department of Psychology, University of Alberta, Edmonton, Alberta,
Canada T6G 2E9. Electronic mail should be sent to: dgrant@psych.ualberta.ca.
|
|

