| "Generalization within classes
and discrimination between classes - this is the essence of concepts"(Keller
& Schoenfeld, 1950, p. 155). |
II. Exemplar View
The universality and simplicity of
generalization and discrimination has led many theorists to the most elementary
level of categorization, i.e. behavioral analysis (e.g. Wasserman &
Astley, 1994). According to behavioral analysis, which is based on the
principles of conditioning, classes represent sets of stimuli with identical
functions. These sets of stimuli are unified, not by a rule, a common feature,
or an abstract representation, but by a common psychologically significant
consequence (Skinner 1935). In the terminology of human categorization
theorists, exemplar models predict categorization in such a manner.
Exemplar models are the most parsimonious
models of categorization in terms of the underlying associative mechanism
(see Chase & Heinemann (2001) for more on exemplar models and an actual working example).
Proponents of exemplar models assume that intact stimuli are stored in
memory, and that classification or recognition is determined by the degree
of similarity between a stimulus and the stored exemplars. Simple generalization
effects explain correct classification of novel (previously unseen) instances
of categories. The tendency to respond to instances to which an individual
has already been exposed will generalize to a similar, novel stimulus when
it is first presented. This means that only the item information is used
for classification decisions, and that categorization relies on the comparison
of a new stimulus with known exemplars of the category. Therefore, generalization
gradients have to be anchored to individual stimuli, and the individual
exemplars have to maintain their memorial integrity even if the number
of experiences relevant to a category is increased dramatically.
There are, however, a number of problems
with the exemplar theory of categorization. Proponents of such theories
cannot agree as to: a) how many exemplars can be stored or retrieved for
comparison, and b) how similarity can be computed so as to ensure that
responding generalizes only to those instances of the same category. The
first problem is not only a matter of memory capacity, but also of feasibility.
Categorization, according to exemplar based processing, is determined by
the number of exemplars to be remembered. "Therefore, any exemplar model
which assumes that all experienced exemplars are remembered may not apply
to pigeon categorization when birds are trained with large numbers of non-repeating
stimuli" (Bhatt et al., 1988). This criticism of the exemplar model
can be overcome by assuming that categorization is based on a small subset
of the total number of stimuli, or that specific retrieval rules act to
determine which patterns are most likely to be accessed. The same is also
true of novel stimuli presented in a generalization test. The average distance
model (Reed, 1972), for example, assumes that the classification process
involves the computation of similarity distances between the new pattern
and all known members of a category. According to this extreme variant
of the exemplar model of categorization, the subject stores all experienced
patterns in memory, and retrieves information when a new pattern has to
be classified. Some exemplar models do, however, allow for more abstract
representations (e.g., Medin & Schaffer's context theory, 1978).
One may question the relevance of
artificial tasks for natural categorization. Learning about perceptual
classes in nature by recognizing every instance may sound like an impossible
feat, but it has to be taken seriously. Although the number of possible
pictures of "a tree" or "not a tree" is infinite, the actual number used
in any particular experiment is likely to be finite. A series of experiments
reported by Vaughan and Greene (1984), for example, revealed that the pigeon
has quite striking powers of exemplar learning and retention. In these
frequently cited experiments, eighty pictures of outdoor scenes were arbitrarily
divided into two categories, positive and negative, and shown to three
pigeons daily in different sequences. The only way in which performance
on this task could have improved would be if each pigeon memorized whether
each exemplar was positive or negative, since there was neither a concept
nor a feature rule that distinguished the two categories. Against the odds,
the pigeons were not only able to sort the 80 pictures correctly (with
a probability of discriminating by pure chance well beyond 10-10), but
they also learned to respond correctly to no less than 320 such pictures.
Fersen and Delius (1989) subsequently
obtained even more outstanding results. Their pigeons were capable of discriminating
100 different positive stimuli from a further 625 similar negative stimuli.
Tests conducted with novel stimuli indicated that the birds had not only
memorized the positive stimuli, but also the negative ones. Furthermore,
the birds retained this information over a period of several months, supporting
the view that persistent and capacious memories are not restricted to food-storing
animals, but may reflect a fitness advantage accruing from extensive and
thorough knowledge of the environment.
Given the remarkable capacity of
pigeons for learning specific exemplars, some researchers may tend to judge
it as a sign of dullness, rather than of intelligence. The ability to learn
relational or abstract concepts is more likely to provide evidence of intelligence.
Throughout this chapter, however, I will argue for the need to be cautious
in this respect. Greene (1983) provides an illustrative example of the
overestimation of the pigeon's spontaneous discriminative abilities. Greene
trained pigeons to discriminate between slides on the basis of a "repetition"
concept. However, those birds actually mastered the task by responding
to minute differences between the two "copies" of the slides. In the following
section, I will offer yet another example of exceptional exemplar recognition
by pigeons that were originally trained on an abstract concept. In this
example, the pigeons were required to discriminate between different "chess-board
patterns" according to the presence or absence of vertical symmetry.
Learning
By Rote Instead of Abstracting A Symmetry Concept
The data presented in this section
are from a long-term study of symmetry recognition that involved several
stages and variants (Huber et al., 1999). However, I will confine my report
to a single experiment that provides ample evidence of the pigeons' ability
to switch from learning about the experimenter's abstract  class rule to learning about the more congenial instances. We used in this,
and in all following experiments, members of the lively and robust "Strasser"
strain of pigeon. A total of 16 subjects were trained to discriminate
a set of 20 vertically symmetrical chessboard-patterns (assigned as "positives"
for one group, and as "negatives" for the other group) from a set of 20
patterns without a symmetrical axis (with the reverse contingencies).
class rule to learning about the more congenial instances. We used in this,
and in all following experiments, members of the lively and robust "Strasser"
strain of pigeon. A total of 16 subjects were trained to discriminate
a set of 20 vertically symmetrical chessboard-patterns (assigned as "positives"
for one group, and as "negatives" for the other group) from a set of 20
patterns without a symmetrical axis (with the reverse contingencies).
We used two different types of black-and-white
chessboard patterns in order to test certain models of symmetry recognition.
One type (the "COMPACT" pattern) consisted of a vertical axis with "whiskers"
on both sides. In the case of the symmetrical pattern, the whiskers on
one side of the vertical axis were a mirror image of the whiskers on the
other side.  In the case of the asymmetrical pattern, the whiskers were randomly distributed
across both sides of the axis (Figure
3). The outer contours of all the figures were filled black.
The exemplars of the second type of pattern (the "SCATTERED" pattern) were
much more like chessboard patterns; i.e. the small elements were scattered
across a 6x6 m
In the case of the asymmetrical pattern, the whiskers were randomly distributed
across both sides of the axis (Figure
3). The outer contours of all the figures were filled black.
The exemplars of the second type of pattern (the "SCATTERED" pattern) were
much more like chessboard patterns; i.e. the small elements were scattered
across a 6x6 m atrix
(Figure
4). However, it would detract from the main point of this study
if the differences between these two types of stimuli with respect to the
assessment of symmetry were discussed more thoroughly. Only the striking
difference between the learning curves of the two experimental groups warrants
mentioning this difference.
atrix
(Figure
4). However, it would detract from the main point of this study
if the differences between these two types of stimuli with respect to the
assessment of symmetry were discussed more thoroughly. Only the striking
difference between the learning curves of the two experimental groups warrants
mentioning this difference.
Members of the stimulus classes appeared,
one at a time, on a computer monitor close behind a transparent pecking-key.
The pigeons were required to peck frequently when a positive stimulus appeared
in order to obtain food, and to suppress pecking when a negative stimulus
appeared in order to terminate a non-reinforced trial. This type of go/no-go
procedure (see details in Figure
5 below) was first used in Herrnstein's laboratory (Herrnstein et al,
1976; Herrnstein, 1979; Vaughan & Greene, 1984) and has also been repeatedly
employed in our own laboratory.
|
Figure 5:
The go/no-go successive discrimination procedure
A
standard training session--which is 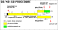 administered once a day, five times
a week--consists of 40 trials (elementary training unit). At the beginning
of a trial a stimulus is presented directly behind the pecking key. During
the first 10 s of a trial, pecks emitted onto the response key are counted
but have no scheduled consequences. Only these responses enter into the
data analysis. Following this 10-s period, and a further 10-s variable
interval (VI) with a range of 1-20 s, subjects enter the consequential
phase of the experiment. If the stimulus is positive, the first response
to occur within 2 s of the previous response results in reinforcement being
made available for a 3-s duration. If a negative stimulus is presented,
the trial is terminated if the pigeon is withholds pecking for 8 sec
during the consequential phase. If, however, the bird pecks continuously
during the presentation of the negative picture, following the timing-out
of the VI schedule, the picture presentation remains active. After trial
termination, an inter-trial-interval (ITI) of 3 s follows, during which
time the monitor is dark. The sequence of positive and negative trials
follows a pseudo-random schedule. No more than 4 positive or 3 negative
trials can occur in a row and the first trial is always positive. administered once a day, five times
a week--consists of 40 trials (elementary training unit). At the beginning
of a trial a stimulus is presented directly behind the pecking key. During
the first 10 s of a trial, pecks emitted onto the response key are counted
but have no scheduled consequences. Only these responses enter into the
data analysis. Following this 10-s period, and a further 10-s variable
interval (VI) with a range of 1-20 s, subjects enter the consequential
phase of the experiment. If the stimulus is positive, the first response
to occur within 2 s of the previous response results in reinforcement being
made available for a 3-s duration. If a negative stimulus is presented,
the trial is terminated if the pigeon is withholds pecking for 8 sec
during the consequential phase. If, however, the bird pecks continuously
during the presentation of the negative picture, following the timing-out
of the VI schedule, the picture presentation remains active. After trial
termination, an inter-trial-interval (ITI) of 3 s follows, during which
time the monitor is dark. The sequence of positive and negative trials
follows a pseudo-random schedule. No more than 4 positive or 3 negative
trials can occur in a row and the first trial is always positive. |
If pigeons learn, this procedure results
in a high rate of responding to patterns identified as positive, and a
low or zero-rate of responding to patterns identified as negative. In trials
pigeons are uncertain as to what group a stimulus belongs, they peck at
an intermediate rate. Learning speed and accuracy are measured using Herrnstein's
rank-order statistic, rho.
Plotting
the learning curves of the two  groups in a single figure (Figure
6) reveals a surprising pattern of results. The investigation of symmetry
conceptualization was overshadowed by a strong effect of stimulus type.
Pigeons that were presented with the compact patterns experienced considerable
difficulty when discriminating between symmetrical and asymmetrical stimuli.
On the other hand, pigeons presented with the scattered figures showed
substantial discriminative abilities. This result is surprising mainly
with respect to the abstraction of the symmetry rule. If this rule is nothing
more than a logical device, then no difference between the types of stimuli
should have occurred. For example, humans informed about the underlying
concept would have no difficulty in sorting the figures in both cases.
If its acquisition, however, is bound to particular visual aspects of the
figures, as many pattern recognition theories suggest (e.g. Osorio, 1996),
then one may wonder whether we should continue to speak about abstract
concepts at all. In any case, generalization tests involving the presentation
of novel stimuli should allow us to determine whether the successful subjects
learned the task solely by memorizing the specific training exemplars.
groups in a single figure (Figure
6) reveals a surprising pattern of results. The investigation of symmetry
conceptualization was overshadowed by a strong effect of stimulus type.
Pigeons that were presented with the compact patterns experienced considerable
difficulty when discriminating between symmetrical and asymmetrical stimuli.
On the other hand, pigeons presented with the scattered figures showed
substantial discriminative abilities. This result is surprising mainly
with respect to the abstraction of the symmetry rule. If this rule is nothing
more than a logical device, then no difference between the types of stimuli
should have occurred. For example, humans informed about the underlying
concept would have no difficulty in sorting the figures in both cases.
If its acquisition, however, is bound to particular visual aspects of the
figures, as many pattern recognition theories suggest (e.g. Osorio, 1996),
then one may wonder whether we should continue to speak about abstract
concepts at all. In any case, generalization tests involving the presentation
of novel stimuli should allow us to determine whether the successful subjects
learned the task solely by memorizing the specific training exemplars.
Such an endeavor requires the presentation
of specifically selected test patterns, rather than the presentation of
previously unseen instances of the training sets. In principle, the ability
to capitalize on abstract relations, such as symmetry, can expand the class
boundary by an unlimited amount, since boundaries will no longer be restricted
by absolute class characteristics. Virtually any picture or object that
is bilaterally symmetrical should be correctly classified when it is first
encountered. Generalization then becomes a matter of inference, rather
than of perceptual similarity. If, on the other hand, common features are
involved, then transfer should occur along this dimension. Generalization
will then become a matter of similarity. Finally, learning the training
stimuli by rote in a photographic, integrative manner should severely impair
generalization performance. Even minute changes in the idiosyncratic structure
of memorized stimuli should result in a sharp trade-off. Generalization
should then become a matter of indiscriminability.
In the type of transfer tests that
we have chosen, we were able to determine the pigeons' categorization strategy
from their unforced choice of novel stimuli, which were selected because
of their similarity to the training patterns. In the first transfer test
we simply used novel exemplars from homologous sets. Twenty novel compact
figures and 20 novel scattered ones were interspersed in further training
sessions  (see stimuli in Figure
7). We then examined whether transfer was limited to the training set
by exchanging the test stimuli, e.g. showing the COMPACT group the scattered
figures and vice versa. A negative result here would imply that transfer
was dependent on absolute class characteristics. Even if the pigeons did
master the transfer test, it would not provide unequivocal evidence of
the possession of a pure abstract symmetry concept. Therefore, a more precise
assessment of whether performance was contaminated by remembered stimulus
aspects was sought using a final test that involved modified training patterns
(see stimuli in Figure
8). For each group, we modified five symmetrical training patterns
in six steps; three of which generated asymmetric patterns. These steps
corresponded to various amounts of change
(see stimuli in Figure
7). We then examined whether transfer was limited to the training set
by exchanging the test stimuli, e.g. showing the COMPACT group the scattered
figures and vice versa. A negative result here would imply that transfer
was dependent on absolute class characteristics. Even if the pigeons did
master the transfer test, it would not provide unequivocal evidence of
the possession of a pure abstract symmetry concept. Therefore, a more precise
assessment of whether performance was contaminated by remembered stimulus
aspects was sought using a final test that involved modified training patterns
(see stimuli in Figure
8). For each group, we modified five symmetrical training patterns
in six steps; three of which generated asymmetric patterns. These steps
corresponded to various amounts of change  (either two, four, or six square elements have been displaced, omitted,
or added). Thus, transfer behavior could vary according to either a) degree
of symmetry, or b) amount of change. If it were found that the birds were
influenced by a), then this would provide evidence of symmetry conceptualization.
Whereas an effect of b) would indicate that transfer was the result of
similarity to memorized training patterns.
(either two, four, or six square elements have been displaced, omitted,
or added). Thus, transfer behavior could vary according to either a) degree
of symmetry, or b) amount of change. If it were found that the birds were
influenced by a), then this would provide evidence of symmetry conceptualization.
Whereas an effect of b) would indicate that transfer was the result of
similarity to memorized training patterns.
The first two tests, which involved
the presentation of test stimuli from the birds' own training class as
well as from a foreign training class, revealed that only among the successful
birds of Group SCATTER did performance generalize to novel instances. However,
even this successful transfer was restricted to those instances belonging
to the birds' own training class (SCATTER), but not to the foreign class
(COMPACT). That transfer performance was severely bound to common absolute
stimulus characteristics was shown more directly by "tacitly" presenting
"ambiguous" figures, or stimuli that closely resembled (symmetric) training
patterns but whose symmetry content had now changed (those three modifications
of each of five originally symmetric figures that now became asymmetric; see
Figure 8).
T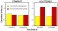 his
forced choice test revealed that generalization was solely the result of
similarity rather than the possession of a symmetry concept. Figure
9 shows that all test stimuli were judged as similar to symmetric training
stimuli (little deviation from symmetrical training stimuli, yellow bars),
but as dissimilar to asymmetric training stimuli (large deviation from
symmetrical training stimuli, red bars), regardless of their own symmetry
type. Thus, irrespective of whether a test stimulus was symmetric or
asymmetric, it was classified according to its similarity to the symmetric
training stimulus it was derived from. Transfer was a matter of similarity
and not of symmetry. Pigeons generalized, if at all, very conservatively
(i.e., to only a small range around the stored templates, much like a cross-correlation-based
template-matching system, Cerella, 1990).
his
forced choice test revealed that generalization was solely the result of
similarity rather than the possession of a symmetry concept. Figure
9 shows that all test stimuli were judged as similar to symmetric training
stimuli (little deviation from symmetrical training stimuli, yellow bars),
but as dissimilar to asymmetric training stimuli (large deviation from
symmetrical training stimuli, red bars), regardless of their own symmetry
type. Thus, irrespective of whether a test stimulus was symmetric or
asymmetric, it was classified according to its similarity to the symmetric
training stimulus it was derived from. Transfer was a matter of similarity
and not of symmetry. Pigeons generalized, if at all, very conservatively
(i.e., to only a small range around the stored templates, much like a cross-correlation-based
template-matching system, Cerella, 1990).
The difference in the ability of
the two groups of pigeons to store their respective training instances
remains to be explained. If pigeons are living "photo-cameras", with large
films that store the pictures they are presented without any further decomposition
processes occurring, then such huge learning differences should not have
occurred. Clearly, if one is only interested in models of categorization
or conceptualization, then one can omit further discussion of this finding,
or leave it to theories of pattern recognition and memory (as was done
predominantly in the human literature on template matching). Those interested
in the basic mechanisms underlying categorization in animals, however,
require a more rigorous investigation of how pictures are processed in
the animal's brain. This means that we will have to enter into a discussion
of how visual objects or scenes are represented in the brain, and what
aspects of these objects or scenes are used for discrimination or categorization
tasks.
|
Exemplar-based
categorization of birds versus mammals
A very clear support for the exemplar
view of visual categorization was found in an experiment by Cook et
al. (1990) in which pigeons were required to learn a discrimination between
line drawings depicting naturally occurring objects, namely birds and mammals.
The primary evidence for this came from the uniform rates of discrimination
acquisition among groups that were trained either with the S+ and S- stimuli
unified by a category (the true categorization condition) or with reinforcement
being uncorrelated with category membership (the pseudoconcept condition).
Further evidence for learning each exemplar separately came from the significant
facilitation of acquisition by using only 5 instances per category instead
of 35 instances per category. This difference in learning speed can
be explained by a lack of within-category generalization, an important
facet of relational or featural learning. Finally, the fact that transfer
to novel category instances was determined not by the typicality of the
pictures ('good' or 'poor' exemplars as assessed by human raters) but by
the specific nature of the exemplars used during training is also in line
with the exemplar view of categorization.
An important facet of this experiment
is the successful specification of those aspects of the stimulus array
that entered into the pigeons' memory. Using specific test variants of
the pictures, the authors found that not the entire picture was stored,
but only the animal figures themselves. From these figures, some features
were not controlling the pigeons' classification behavior, like the 90
degree rotation or the reflection about the vertical axis. It seems as
if the birds had selectively attended to specific aspects of the stimulus
array and that they had decomposed the pictures into at least figure and
ground. Such analytic processes, however, are perfectly in line with a
more feature-based account of categorization and this apparent theoretical
incompatibility points to a fundamental problem of perceptual learning:
How do information-processing systems make determinations of similarity
(Blough, 2001; Lea, 1984;
Shepard, 1987)? |
In principle, we may distinguish between
item-specific and category-specific aspects of the stimuli. The first ones
are needed to discriminate between instances of the same class; the second
ones are needed to discriminate between instances of different classes.
Differential within- versus between-class generalization is commonly considered
as a key feature of visual categorization in animals (Wasserman & Astley,
1994). If no category-specific information is available as a kind of relational
information about common properties of a category, then classification
learning will be restricted to learning about each individual stimulus separately
and distinctly. If it were available, learning to use it will significantly
facilitate classification, both in terms of acquisition and transfer to
novel instances. The facilitation of classification is due to intra-class
generalization, that is transfer to novel instances of the same class is
mediated by generalization or item similarity to the previously learned
exemplars. The classic procedure to disclose learning about category-specific
features is the pseudoconcept task which involves the arbitrary assignment
of category (for example "fish", Herrnstein & de Villiers, 1980) and
non-category exemplars ("non-fish" in this example) to positive and negative
classes, respectively. Lea and Ryan (1990) called this the "perverse
pseudoconcept task" in order to distinguish it from the alternative
"random pseudoconcept task" in which no 'concept' exists within
the stimulus set).
A number of categorization experiments
involving such pseudoconcept tests have successfully shown that pigeons
learn about category-specific information in contrast or in addition to
item-specific information (Herrnstein & de Villiers, 1980; Edwards,
& Honig, 1987; Pearce, 1988; Wasserman, Kiedinger & Bhatt, 1988).
On the other hand, using a similar procedure, some authors have found differences in acquisition rate and concluded that the animals achieved
a discrimination by learning only about the specific stimuli in the acquisition
phase (Vaughan & Greene, 1983; Schrier, Angarella & Povar, 1984;
Cook, Wright & Kendrick, 1990).
A very reasonable suggestion brought
forward by Cook et al. (1990, also see sidebar) is that categorical discriminations
may consist of two phases, a "stimulus learning" and a "concept" phase.
The first would involve only learning about the exemplars by attending
to the item-specific information that distinguishes them from all other
stimuli. The second phase, in which category-specific information is extracted,
may then follow. In cases in which it doesn't, as in our symmetry experiment
above, true open-ended categorization as a means of minimizing memory requirements
is not achieved. It is fair to say, that absolute, data-driven, exemplar-based
strategies of categorization are a plausible alternative to more relational
and analytic processing mechanisms, especially from an engineering point
of view (Cook et al., 1990). However, the relevance for pigeons and other
animals in the wild remains to be specified (see an extensive discussion
of this issue in Huber, 1995, 1999).
Next Section: Feature
Learning
