III. Feature Learning
The term "feature" refers to any
elementary property of a distal stimulus that is psychologically processed--an
"atom of cognition". It is usually used to refer to the fixed properties
of a stimulus, which are the most primitive or the lowest building blocks
of object recognition and categorization (e.g. Bruner, Goodnow & Austin,
1956). Although the advantages of a feature learning over a template approach
to categorization (e.g. Ullman, 1989) are already available in terms of
straightforward models of similarity (e.g. Shepard, 1987; Blough,
2001) and of structured hierarchical representations, some
would argue in favor of more flexible features (Steele, 1990; Schyns, Goldman
& Thibaut, 1998). While a very large set of object descriptions and
categories can be generated from a finite set of elements and combination
rules, the fixed feature approach is limited to the possible combinations
of the feature set.
Feature Analysis
Versus Feature Learning
Feature analysis refers to the fact
that a subject or species enters into a problem with a fixed set of features,
to which it is sensitive. It will consequently fail the task if the
categories to which it is exposed are not separable along these feature
dimensions. If, furthermore, combinatorial power is also limited, then
the animal will be required to classify objects in the natural environment
by fixing on some specific, single feature. This ability is a natural example
of a feature analysis system, and also exemplifies the power of such a
simple strategy (Uexkuell, 1939; see also Herrnstein, 1985).
It is easy to imagine occasions on
which features, not originally present in the system, are useful for distinguishing
between important categories in the world that confront the organism (Schyns,
Goldman & Thibaut, 1998). Feature learning would be able to tailor
the animal's feature repertoire to the demands of categorization. The set
of features used and/or the salience of these features could be continuously
modified during the course of categorization. Such a flexible system would
be able to adapt to a changing environment, especially in those situations
in which it would be unrealistic to think that a system comes fully equipped
to deal with them appropriately. Artificial tasks created in the laboratory
provide a straightforward way of testing this ability.
A number of categorization experiments
with animals that can be described in terms of feature theory have already
been conducted. This is particularly apparent in those cases in which the
task involved artificial stimuli devoid of the richness of natural scenes.
Researchers have attempted to interpret these results by trying to identify
those aspects that controlled the animals' classification performance.
Such post-hoc analyses have been conducted either by correlating the feature
values that the experimenter was attending to with the animals' response
rates, or by submitting the data to more sophisticated statistical procedures,
such as cluster analysis and multidimensional scaling (see
Blough, 2001). The letters of the alphabet are stimuli that have been
used repeatedly in such experiments (Blough, 1985; Lea and Ryan, 1983;
Morgan, Fitch, Holman & Lea, 1976).
An even more illustrative example
of artificial stimuli is the cartoons of the Peanut family used in the
lab of Cerella (1980, see also Cerella, 1982, 1986). The 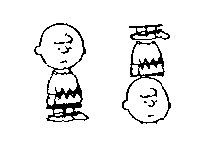 striking result of this frequently cited experiment, was the pigeons' resistance
to deformations of "Charlie Brown" (Figure 10, see also Kirkpatrick,
2001). This finding is interesting because it is hard to reconcile
it with our understanding of template action. If the animal is attending
not to the entire figure, but only to some small component, then generalization
should be preserved even if Charlie Brown is truncated, scrambled, or presented
upside-down. A feature model need only assume that the local aspects of
figures are processed separately and are position-invariant.
striking result of this frequently cited experiment, was the pigeons' resistance
to deformations of "Charlie Brown" (Figure 10, see also Kirkpatrick,
2001). This finding is interesting because it is hard to reconcile
it with our understanding of template action. If the animal is attending
not to the entire figure, but only to some small component, then generalization
should be preserved even if Charlie Brown is truncated, scrambled, or presented
upside-down. A feature model need only assume that the local aspects of
figures are processed separately and are position-invariant.
The strength of any feature model
lies in its ability to explain virtually any categorization behavior, and
in its plausibility in terms of informational economy. Furthermore, successful
generalization can be explained in terms of common features and the associative
strength that they acquire during training, rather than by evoking an obscure
concept or straining the template account. If we accept that the presence
or absence of a certain set of features is responsible for determining
how a subject classifies an individual stimulus, then we need only specify
how these features acquire their influence. If the total number of possible
features belonging to separate classes of stimuli are predetermined, then
one only has to specify whether responding has come under the control of
these features. If this is the case, and a clearly specified feature model
is able to describe the data adequately, then it would be superfluous to
ascribe to pigeons the formation of a concept. Unsurprisingly, several experiments
using this criterion have been successfully conducted (Jitsumori, 1993;
Lea & Ryan, 1983; Fersen & Lea, 1990).
Determining
the Controlling Features
Following this line of analysis,
even those experiments that have been described in terms of concept formation,
might be explained in terms of feature-positive discrimination. For example,
if pigeons receive trials in which photographs containing people signal
food, and trials in which photographs without any people signal the absence
of food. Then those features that are characteristic of people will steadily
acquire associative strength due to their positive reinforcement history.
In contrast, all irrelevant features, i.e. those that are present in both
reinforced and non-reinforced trials, will acquire associative strength
in an erratic manner and in the long term will lose attention. The problem
with this approach, however, is that it cannot specify exactly what features
are involved. The scarcity of information about the precise nature of the
cues controlling the discrimination is endemic for categorization tasks
involving complex stimulus classes (Cook et al., 1990; Fetterman, 1996).
Inspection of the slides alone cannot allow us to determine which dimensions
divide the feature space, especially when photographs of natural scenes
are employed. Those experiments that employ slides of people, trees, bodies
of water, fish, flowers etc are resistant to rigorous tests in terms of
feature theory. This is not only true of the experimenter him- or herself,
but also of the critical reader of such experiments who attempts to reevaluate
the results but is not provided with the full set of pictures used (see
Premack, 1983; Editor's comment: this should change with the increased
use of the web as is evidenced many times in this volume).
One way of determining which aspects
of a complex picture, or class of complex pictures, have acquired control
over the subjects' responding is to analyze the subjects' misclassifications.
Experimenters have sometimes found better performance on those slides that
they would have classified as intermediate to poor with respect to the
saliency of the underlying concept (Herrnstein & De Villiers, 1980;
Schrier & Brady, 1987). This finding suggests that, in these experiments,
the subjects' behavior may have been under the control of a slightly different
concept to that intended by the experimenter. Analysis of those slides
that elicited crude misidentifications, has proven to be especially informative,
since it has evoked rather strong reservations about the conceptual ability
of the subjects under investigation (e.g., D'Amato & Van Sant, 1988).
D'Amato and Van Sant's (1988) monkeys, for example, showed persistent errors
to those slides including patches of red coloration (e.g., a piece of watermelon
in the center of a table). Such features are no doubt irrelevant from the
point of view of the human concept holder, but not from the point of view
of the monkey. The same control of animals' behavior by the background
of photographs was also found in Green's (1983) reevaluation of Herrnstein's
experiments, and in Honig and Stewart's (1988) experiment.
While it is clear that persistent
reliance on the irrelevant features of a target concept is cause for serious
concern about whether animals form concepts at all, it strongly supports
the more parsimonious feature account. Animals do not learn about the defining
features of a concept and neglect all other features that may be present,
but are not necessary properties or characteristics of any member of the
target concept. Rather they learn about any feature that occurs with some
positive probability on trials and are followed by a specific psychological
consequence (e.g. food).
As mentioned above, one severe problem
with the feature account of natural categorization in animals is that the
stimulus aspects controlling the subjects' behavior cannot be completely
and reliably specified. One way to render this task more manageable is
to use stimuli that are poor in detail and describable only in terms of
a few well-understood features. The advantage of such an approach is two-fold.
First, the number of stimulus aspects that might occur with a high probability
can be reasonably reduced. For example, using line drawings can make a
"hidden" feature such as a specific one-sided distribution of a hue or
the amount of background (Schrier & Brady, 1987) unlikely to correlate
with one stimulus class.
Second, the low-dimensional feature
space of artificial stimuli enables the experimenter to arrange stimulus
classes along a linear discriminant function (Nilsson, 1965). In terms
of geometry, this means that the two categories to be discriminated by
the animal occupy different areas in a multidimensional feature space.
Such a clear-cut experimental design, that forces the subject to adopt
the categorization rule or else fail, is achievable only if the stimuli
are constructed by the experimenter.
More rigorous tests of the subjects'
ability to extract the relevant features, and to combine them in a manner
that most closely corresponds to the experimenter's categorization rule,
requires a synthetic approach to categorization. This has been adopted
with considerable success in Stephen Lea's laboratory in Exeter (Lea &
Harrison, 1978; Lea, Lohmann & Ryan, 1993; Lea & Ryan, 1990; Fersen
& Lea, 1990; see also the research of Shimp,
Herbranson, & Fremouw, 2001) The synthetic approach
is an a priori method that involves the construction of artificial concepts
that can be defined by a small number of independent features, each of
which comes with a predetermined probability of occurrence. One experiment
designed according to all the above criteria was conducted as part of my
Ph.D. thesis (Huber, 1991) and published thereafter (Huber & Lenz,
1993).
A Test of the
Linear Feature Model
The two classes of stimuli used in
this experiment could be distinguished on the basis of a "polymorphous
feature rule". Ryle (1949) coined this term in order to describe the nature
of ordinary language concepts. Wittgenstein (1953) argued, in a similar
manner, that natural categories are ill-defined, i.e. they lack any singly
necessary or jointly sufficient attributes of definition. More simply,
the features of natural categories are not present in all exemplars, but
are only characteristic or likely to be class properties. Consequently,
the members of such categories do not have equal status, but are more or
less typical of the category depending on the number of characteristic
features that they possess. In order to approximate this philosophical
account, and to make it manageable for psychological experiments, several
researchers have used an "m-out-of-n rule" (Dennis, Hampton & Lea,
1973; Shepard, Hovland & Jenkins, 1961). According to this rule, membership
information is a quantitative combination of several independent feature
dimensions. Dennis, Hampton & Lea (1973), for example, asked students
at the University of Cambridge to discriminate between artificial sets
of stimuli that were defined by a two-out-of-three rule. Positive stimuli
contained two out of possible three positive feature values, while negative
stimuli contained two out of possible three negative feature values.
Lea and Harrison (1978), using a
similar procedure for the first time with pigeons, reported successful
training of the six training stimuli constructed using the above rule,
and successful generalization to the two test stimuli (which contained
all the positive or all the negative feature values). However, the very
small number of training stimuli used in this experiment casts doubt upon
whether the pigeons really learned about the features at all, or whether
they just learned about the patterns in their entirety. Therefore, in my
own experiments I used 62 stimuli, which I believed would increase the
probability of feature learning. Simple line drawings were created that
resembled, to the human observer, human faces (called "Brunswik faces").
Three examples of these patterns are depicted below (Figure 11).
Egon
Brunswik originally used these types of stimuli in an investigation of human  expressions (Brunswik & Reiter, 1937). Many years later, Reed (1972)
used them to investigate human visual categorization. We created them on
the computer and presented them on black-and-white slides with luminous
contours and dark backgrounds. The schematic faces consisted of four facial
features that could be assigned one of three values. The four facial features
were distance between the eyes (e), height of the forehead (brow, b), length
of the nose (n), and position of the mouth (chin, c). All four features
were assigned values of -1, 0, or +1 (arbitrary values that correspond
to equally large displacements). Using this method, it is possible to generate
81 (34) different faces. The next step involved the arbitrary assignment
of the feature values to the categories. The pattern containing a short
distance between the eyes, a high brow (a low eye position), a short nose,
and a high chin (a high mouth position) was designated as the prototypical
negative pattern [e-b-n-c-] (left figure); while the pattern containing
a wide distance between the eyes, a small brow (a high eye position), a
long nose, and a small chin (a low mouth position) was designated as the
prototypical positive pattern [e+b+n+c+] (right figure). The intermediate
figure is depicted in the center.
expressions (Brunswik & Reiter, 1937). Many years later, Reed (1972)
used them to investigate human visual categorization. We created them on
the computer and presented them on black-and-white slides with luminous
contours and dark backgrounds. The schematic faces consisted of four facial
features that could be assigned one of three values. The four facial features
were distance between the eyes (e), height of the forehead (brow, b), length
of the nose (n), and position of the mouth (chin, c). All four features
were assigned values of -1, 0, or +1 (arbitrary values that correspond
to equally large displacements). Using this method, it is possible to generate
81 (34) different faces. The next step involved the arbitrary assignment
of the feature values to the categories. The pattern containing a short
distance between the eyes, a high brow (a low eye position), a short nose,
and a high chin (a high mouth position) was designated as the prototypical
negative pattern [e-b-n-c-] (left figure); while the pattern containing
a wide distance between the eyes, a small brow (a high eye position), a
long nose, and a small chin (a low mouth position) was designated as the
prototypical positive pattern [e+b+n+c+] (right figure). The intermediate
figure is depicted in the center.
According to the m-out-of-n principle,
the categorization rule can be defined as the quantitative combination
(or sum) of feature values, plus a threshold criterion that separates the
two categories. By this definition, each pattern with a negative feature
sum (n=31) belongs to one category (NEG), while each pattern with a positive
feature sum (n=31) belongs to the other category (POS). (The 19 patterns
with a feat ure
sum of zero were omitted from the experiment.) Inspection of these stimuli
(Figure 12) reveals
that no single feature dimension alone is a perfectly reliable cue, but
has an equal, low value of membership predictability. In other words, attending
to eye distance alone can never provide sufficient information to determine
trial outcome. The same is also true of the feature values. Negative values
are correlated with category NEG and positive values with category POS,
but only weakly (p = 0.55). Furthermore, even the opposite value (negative
for category POS, and positive for category NEG) occurs occasionally (p
= 0.13). Finally, feature values of zero occur equally often in both
categories (p = 0.32).
ure
sum of zero were omitted from the experiment.) Inspection of these stimuli
(Figure 12) reveals
that no single feature dimension alone is a perfectly reliable cue, but
has an equal, low value of membership predictability. In other words, attending
to eye distance alone can never provide sufficient information to determine
trial outcome. The same is also true of the feature values. Negative values
are correlated with category NEG and positive values with category POS,
but only weakly (p = 0.55). Furthermore, even the opposite value (negative
for category POS, and positive for category NEG) occurs occasionally (p
= 0.13). Finally, feature values of zero occur equally often in both
categories (p = 0.32).
As predicted by Wittgenstein and
Ryle, the instances of a polymorphous class are not equally valid members
of the categories to which they belong. Typicality of category membership
varies along the summary dimension; one pattern in each class (with an
absolute feature sum of four) is a perfect category member, patterns with
an absolute sum of three are very typical, those with an absolute sum of
two are intermediate, and those with an absolute sum of one are poor members.
Therefore, the only way to classify
the stimuli exactly is to respond to their "feature summary". In order
for successful categorization to occur we predicted that the pigeons must
be able to:
-
Extract information about, or attend
to, all four features of class membership
-
Equalize the weights given these features (or resist selective
attention)
-
Combine this feature information in an additive manner.
Based on these predictions, the model
was supported in two ways. Pigeons were trained on all 62 stimuli, but
in a stepwise manner in order to investigate how stimuli introduced at
later stages are classified (a form of generalization test). Afterwards,
several independent observers assessed the veridicality of the above criteria,
i.e. whether selective attention to specific facial features varied, and
whether the features were combined additively or in a more integrative
manner.
The pigeons' learning performance
was surprisingly good. After only three weeks of training, which consisted
of three stages that involved an increasing number of stimuli (first 10,
then 20, and finally 60), the three experimental subjects were able to perfectly
divide all stimuli into the categories defined by the experimenter.
Inspection of the learning curves (Figure
13) reveals that some disruption occurred following the shift from
the first to the second training stage. However, the fact that these errors
occurred with an equal frequency during the presentation of the already
known stimuli and the novel ones, led us to reject pattern learning effects.
From session 13 onwards, when the number of training stimuli was increased
from 20 to 60, we found highly accurate initial responses to novel stimuli.
Both birds that continued discriminating (unfortunately pigeon, st3, died)
demonstrated almost perfect categorization abilities. The discrimination
ratios for these two birds, averaged across sessions 15 - 19, were 0.96
and 0.98. This was achieved by pecking between 2.0 - 3.6 times more frequently
to positive stimuli than to negative ones.
Even more informative than the learning
performance of the birds, was the analysis of the pecking rates directed
at individual instances of the categories. According to prediction 1),
all four facial features should have exerted equal control over the birds'
responding. To test this prediction we applied a method already used by
Lea and Ryan (1983). Pecking rates to all 60 stimuli (i.e., mean number
of pecks to each stimulus across all sessions of stage 3) were subjected
to a multi-factor analysis of variance, conducted for each bird separately.
This confirmed that all features had a significant effect on the variance
of pecks across stimuli.
A ccording
to prediction 2) the birds should have paid equal attention to all four
features, i.e. pecking to the features presented with the same attributes
should have been equivalent. It can be seen that plotting the distribution
of pecking rates as a function of feature value during the third training
stage (Figure 14)
fulfilled this criterion.
ccording
to prediction 2) the birds should have paid equal attention to all four
features, i.e. pecking to the features presented with the same attributes
should have been equivalent. It can be seen that plotting the distribution
of pecking rates as a function of feature value during the third training
stage (Figure 14)
fulfilled this criterion.
Finally, according to prediction
3), combining the feature values in an additive manner should produce an
orderly relationship between the sum of the feature values of a  particular face and the rate of responding that it elicited. Although not
achieved in every instance, when we averaged across all stimuli we found
a remarkably orderly correlation between pecking rate and feature sum (Figure
15), sufficient to support a linear feature model (see sidebar below).
particular face and the rate of responding that it elicited. Although not
achieved in every instance, when we averaged across all stimuli we found
a remarkably orderly correlation between pecking rate and feature sum (Figure
15), sufficient to support a linear feature model (see sidebar below).
|
A quantitative
examination of the pigeons' classification behavior
One feature of these results is that
they can easily be interpreted in terms of learning theory. Consider, for
example, one well-known feature model of discrimination. The Rescorla-Wagner
(1972) model predicts that each feature value will gain positive associative
strength during the course of learning, to the extent that it occurs on
positive trials, and vice versa for feature values occurring on negative
trials. We saw earlier that the positive feature value (+1) is correlated
with positive patterns, and that the negative feature value (-1) is correlated
with negative ones. Hence, while positive features will acquire positive
associative strength, negative features will acquire negative associative
strength. Because this model is a form of independent cue model (Reed,
1972), the strength of responding during presentation of a compound stimulus
is determined by a linear combination (sum) of the associative strengths
of its components.
An attempt to quantitatively describe
the classification performance of the experimental subjects is involved
in Huber and Lenz (1993). The R2-values of the multiple regression
functions that involved the four predictor variables (i.e., the values
of the four feature dimensions) exceed, for both birds, the 80% level.
By direct application of the variable weighted linear feature model (Lea
& Ryan, 1990), we can describe the discrimination behavior of the two
pigeons by means of a simple linear equation:
pecks = k + b1e + b2b
+ b3n + b4c.
k is a constant for pecking;
e,
b,
n,
and c are the feature values; and b1-b4 are the weightings of the
feature dimensions. A multiple regression analysis of the data from Bird
st1 yield the following equation:
pecks(st1) = 10.69 + 3.17e
+ 3.51b + 3.26n + 4.97c
For Bird ht1 the equation reads:
pecks(ht1) = 8.8 + 1.66e
+ 2.23b + 1.64n + 1.95c.
|
However, this may not be the whole story.
Although it may seem like I am splitting hairs, the factor responsible
for the remaining proportion of variance in the pigeons' responding remains
to be identified. Look at the following modified Figure 15 in the next
figure (see Figure
16). Especially for Bird ht1 it is easy to recognize that the six points
representing pecking rate as a function of feature sum are not located
precisely on the regression line. If we combine the three points of each
category, then a more or less sharp crack appears. It would appear that
the pigeons have gathered additional information in order to divide the
class even more efficiently than would have been possible if only the summary
information was being used. Statistical evidence for this assumption can
be obtained by inserting an additional factor into the multiple regression
analysis with the value of one for positive stimuli and zero for negative
ones (see also Lea & Ryan, 1983). For one pigeon (ht1), this additional
independent variable that we may call the "category variable" accounted
for a significant proportion of the variance in pecking frequency.
What exactly is the source of the
additional information that the birds were using? One possibility that
we have considered is the use of Gestalt information, extracted from any
relationships between the four facial features. However, the analysis of
variance revealed that there was no interaction between the facial features.
Therefore, another possibility is that learning about the specific configurations
of the exemplars contaminated feature learning. This is not implausible
if one considers that some faces, particularly those already presented
during the first and second training stages, were shown up to 53 times.
The fact that the other pigeons' performance was only marginally affected
by the additional information led us to continue interpreting the results
of this experiment in terms of a feature account.
Elemental
Versus Configural Theories of Conditioning
Some authors are not so confident.
One particularly interesting explanation of our results is in terms of
configural theory (e.g. Pearce, 1987, 1994). According to configural theory,
which is an alternative to an elemental theory of conditioning, "if
a compound stimulus is presented for conditioning, discrimination of categorization,
then a configural representation will be formed of the entire pattern of
stimulation. This representation will then enter into a single association
with the outcome of the trial" (Pearce, 1997, p. 116). The important
point of this statement is that generalization to novel stimuli occurs,
not along separate feature dimensions, but along the distance dimension
on which configurations of stimuli, rather than their feature values, are
located. Applied to categorization problems, stimuli that are repeatedly
paired with food during training will acquire positive associative strength,
and this excitation will generalize to any other similar stimuli presented
for the first time during a transfer test. The same is also true of negative
training stimuli; i.e. inhibition will generalize to other similar test
stimuli. "The interaction of these sources of excitation and inhibition
will leave the new stimulus with a net level of excitation, and it will
elicit a response appropriate to the category to which it belongs"
(Pearce, 1997, p. 122).
It is not difficult to recognize
that there is a close relationship between the configural theory of conditioning
and the exemplar view of categorization. In fact, generalization from stored
intact stimulus compounds during transfer tests has been proposed not only
by Pearce and collaborators (e.g. Pearce, 1988, 1989, 1991; Aydin &
Pearce, 1994), but also by Astley & Wasserman (1992). Whether this
theory has any relevance for the results reported above remains to be specified.
At first sight, the remarkable difference in responding to members of the
same category, (e.g. to members with an absolute sum of three, and to members
with an absolute sum of one), might seem incompatible with the exemplar
view. However, as Pearce (1997) has convincingly argued, faces with a net
sum of +1 are much more similar to faces with a net sum of-1, than to faces
with a net sum of +3. Hence inhibition will be more likely to generalize
to these negative stimuli. Faster responding to a +3 face is therefore
a result of peak shift effects (see also Spence's (1937) theory of discrimination
learning).
As this simple experiment involving
three pigeons categorizing Brunswik faces has demonstrated, it is rather
difficult to disentangle feature theories from exemplar theories in terms
of the general predictions that they make. Despite this difficulty, I attempted
such an endeavor by presenting a specific test stimulus during categorization
training.
The two theories make radically different
predictions about responding to this test stimulus. The test, which followed
the rationale of Medin & Schaffer (1978), was intended to provide support
for context theory (the first formal application of the exemplar theory).
Immediately after the second training phase, five test sessions were administered
that consisted of the presentation of the training stimuli from the second
training phase. However, one specific test stimulus was presented twice
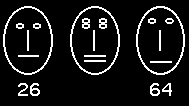 instead of two arbitrarily selected
training stimuli. This specific test stimulus (labeled "26") had a net
sum of -1 and was selected because it was more similar to pattern "64",
a stimulus of the positive class with the net sum of +1, than to any member
of the own, negative training class presented so far (see both stimuli
and the degree of matching below). If--according to the exemplar view--the
initial responding to this test stimulus is determined by its similarity
to the previously experienced training stimuli, hence--at least in part--to
Stimulus "64" which is most similar to it, then we would expect a high
rate of responding. Feature theory, on the other hand, would predict responding
as gingerly as to other faces with a sum of -1.
instead of two arbitrarily selected
training stimuli. This specific test stimulus (labeled "26") had a net
sum of -1 and was selected because it was more similar to pattern "64",
a stimulus of the positive class with the net sum of +1, than to any member
of the own, negative training class presented so far (see both stimuli
and the degree of matching below). If--according to the exemplar view--the
initial responding to this test stimulus is determined by its similarity
to the previously experienced training stimuli, hence--at least in part--to
Stimulus "64" which is most similar to it, then we would expect a high
rate of responding. Feature theory, on the other hand, would predict responding
as gingerly as to other faces with a sum of -1.
Stimulus "26" was presented
10 times and followed by a neutral trial outcome: the termination of the
trial after 10 sec. The average number of pecks elicited by this "ambiguous"
pattern was 2.80, 3.00, and 3.25 for Birds ht1, st1, and st3 respectively.
Pecking during the presentation of the comparison stimulus "64" varied
between 11.25 and 13.50. Furthermore, there was no single case during all
five test sessions, in which a positive stimulus elicited fewer than 10
pecks (pecking to negative stimuli varied between one and 10 pecks). Therefore,
it would appear that all three pigeons recognized Face "26" as signaling
the absence of food--a finding which is in full accordance with feature
theory, but in sharp contrast to the predictions of the exemplar view.
Unsurprisingly, researchers that
favor an exemplar account of categorization over a feature learning account,
will not be sufficiently convinced by this single finding. It is admittedly
difficult to derive any firm conclusions concerning the relative merits
of exemplar and feature theory. Instead of training legions of pigeons
in order to test different versions of the available theories, we can change
to a radically different approach.
Computer simulations based on connectionist
or neural networks (e.g. Gluck and Bower, 1988; Gluck, 1991; Shanks, 1991)
have proven to be particularly useful in this respect. Given that both
elemental and configural theories are already available in the form of
computer models, we only have to feed our data into the different variants.
Because the data from three pigeons seemed to be rather poor for such modeling,
Renate Lenz trained three further pigeons on the original task. The models
were then compared by feeding into the computer the stimuli (or their respective
feature values) in exactly the same order as they were presented to the
pigeons during the various training stages.
Before evaluating the main results
of these simulations, it is worth noting that the performance of the five
pigeons used in these simulations varied in two respects. First with respect
to selective attention during the early training phases, and second with
respect to learning speed. Consequently, the degree of fit of the various
models depends on which subjects were being compared. More specifically,
in all those variants of the models in which we included a variable for
selective attention, the proportion of variance accounted for differed
considerably. In order to examine the data in more detail, they were subjected
to linear regression analyses, using several different combinations of
the predictor variables.
 A
summary of the percentage of variance explained by the linear regression
analyses of the predictions of the different models, together with the
empirical data for each bird is shown in Table
1. The original version of the configural cue model does not account
for selective attention. If the pure version of this model is compared
with the predictions of a feature model, which does not include selective
attention, and feature sum is used as a predictor of pecking behavior,
then a considerable difference is found for fast and slow learning pigeons.
In the case of the two fastest learners (the first two subjects), the feature
sum explains the data significantly better than the configural cue model.
In the case of the two slowest learners, it is the other way round.
A
summary of the percentage of variance explained by the linear regression
analyses of the predictions of the different models, together with the
empirical data for each bird is shown in Table
1. The original version of the configural cue model does not account
for selective attention. If the pure version of this model is compared
with the predictions of a feature model, which does not include selective
attention, and feature sum is used as a predictor of pecking behavior,
then a considerable difference is found for fast and slow learning pigeons.
In the case of the two fastest learners (the first two subjects), the feature
sum explains the data significantly better than the configural cue model.
In the case of the two slowest learners, it is the other way round.
When individual attention weights
were included in the configural cue model, the comparison revealed that
the variably weighted linear feature model, represented here as multiple
regression analysis, is superior for all the fastest learning birds (P1,
P2, P3). The variably weighted linear feature model also revealed better
results for Bird P4. Only for Bird P5, were both models equally good predictors.
However, the predictions of the configural cue model can be further improved
by assuming a higher degree of similarity between the stimuli. This is
true especially in the case of fastest learners P1, P2 and P3. Nevertheless,
for P1 and P3, the variably weighted linear feature model still showed
better results. For the slow learners, P4 and P5, this modification provided
no improvement. Finally, in order to extend the generality of our findings,
we added a further exemplar model. However, using Kruschke's neuronal network
model ALCOVE (Kruschke 1992), we failed to make any further improvements.
In summary, for all pigeons, the
variably weighted linear feature model explained the greatest percentage
of variance. Nevertheless, there are ways in which the predictions of the
configural cue model could be improved, such that for some pigeons (P2,
P4 and P5) the difference was no longer significant. However, the predictions
of the variably weighted linear feature model can also be further improved
for P2 by including additional variables (e.g., a dummy variable and a
variable that represents the frequency of pattern presentation of the individual
stimuli) in the multiple regression analysis. The fact that for the fast
learners (P1, P2 and P3), the variably weighted linear feature model shows
better results than the configural cue model, indicates that some sort
of feature extraction mechanism was involved. However, there might still
be ways in which to improve the predictions of the configural cue model
for individual pigeons (Pearce, 1994). At this point, however, we do not
know of any straightforward way in which to do so.
In conclusion, we may ask what advantages
can be gained by having two models converging, if the models are simulated
by computers in which the boundaries between the theories dissolve in the
net of (hidden) units. For example, it is questionable as to whether an
exemplar model is turned into a feature model by implicitly using individual
feature weightings for representing the stimuli. Furthermore, there is
little agreement concerning how similarity can be correctly assessed, how
many entire patterns enter into associations, and how reliably the models
can be extended to include natural stimuli. Computing similarity in terms
of common elements may seem, at first, to be relatively straightforward.
It is, nevertheless, unclear as to what should be regarded as an element,
even for simple stimuli such as Brunswik faces. Natural stimuli cannot
be seriously simulated in terms of A+ B+ AB-. If, however, they could be
reduced in this radical way, then we are in the midst of the feature theory.
On the other hand, proposing that features are not perceived and are processed
independently, is by no means an objection to feature theory but only to
one traditional, perhaps out-of-date, version.
Further progress must be made in
order to clarify these points. For the present, however, we may agree with
Pearce (1997) "that the mechanisms that are believed to be responsible
for the way animals solve relatively simple discriminations are also likely
to be responsible for the way they solve complex categorization problems."
Limits of the
Feature Account
Continuing to improve the feature
account of categorization requires that we consider how natural categories
are distributed over the feature space. In our experiment, the categories
consisted of many exemplars, which differed only in terms of a few stimulus
dimensions. The decomposition of these patterns into stimulus components
should thus have been fairly easy and economical. Moreover, the separability
of the feature dimensions in the case of the Brunswik faces should have
facilitated this decomposition. It is likely that, integrative dimensions
will be processed by the visual system in substantially different ways.
Finally, the fact that no single cue or feature alone was sufficient for
differentiating between the categories would have favored a strategy that
combined information from several feature dimensions in order to solve
the classification problem.
If we adopt this point of view, then
it is fair to admit that the above findings cannot be generalized limitlessly.
From the literature on animal concept discrimination, and from our own
data, it is possible to derive at least three important constraints on
the use of this strategy:
-
The number and salience of the relevant
features;
-
The 'naturalness' of the defining features;
-
The distribution of the categories to
be discriminated.
A few years ago, Lea, Lohmann &
Ryan (1993) reported an experiment in which pigeons were confronted with
stylized monochrome drawings that consisted of five relevant feature dimensions.
Although this experiment was similar to ours in many respects, the pigeons
experienced considerable difficulty when spontaneously abstracting these
equally balanced features. Although pigeons may be able to attend to these
features when forced to do so, there may be a limit to the number of features
that can simultaneously control behavior. If too many features are relevant
for the concept rule, then the animal may attend to only a subset of the
relevant features, especially if they are very salient. The failure of
dominant features to control behavior may prevent accurate classifications
from being made. Support for this can be found in an experiment with "faked
pigeons" (Lea & Ryan, 1990), in which wing feature was found to control
the classification performance of the birds much more so than any of the
other relevant features. However, despite these occasional problems, feature
learning is not an unreasonable model of categorization. Fersen and Lea
(1990) and Jitsumori and Yoshihara (1997), for example, showed that pigeons
are more than capable of attending to five mutually orthogonal features
simultaneously, and categorizing successfully despite considerable differences
in the salience of the relevant feature dimensions.Because I will discuss
the difficult and important aspect of 'naturalness' in Section V, it may
suffice here to introduce this aspect by referring to an informative experiment
in this respect (Click
here to see material on Fersen & Lea, 1990).
The third factor that is believed
to interfere with category learning is the structure of the similarity
space. As we ourselves have admitted (Huber & Lenz, 1993), the application
of an m-out-of-n rule for category definition, leads to a similarity structure
that may not be representative of natural situations. Since the four feature
dimensions we used did not correlate with one another, a rather "flat"
structure emerged, with no single stimulus rising above the others. Moreover,
the classes did not deviate from one another to occupy distant regions
in the similarity space. For example, in our experiment each category consisted
of 16 stimuli with an absolute sum of one and 15 with an absolute sum of
greater than one. It is possible that the omission of those 19 patterns
with a feature sum of zero might have facilitated category discrimination,
as proposed by Lea, Lohman and Ryan (1993). I doubt whether our Brunswik
classes represent genuine clusters of relatively similar stimuli.
In more natural situations, ambiguous
category exemplars located near to the category border are much less frequent.
To put it more pictorially, natural categories represent mountain landscapes,
with dense regions around class centers and sparse regions near the boundaries.
The reason for this kind of unequal distribution in the similarity space
is the high correlation between features that are class-typical. Trees,
for example, vary considerably, but some features such as green and leaf,
occur together. This "nested" feature distribution has been nicely illustrated
in an experiment by Jitsumori (1993) in which pigeons were shown to attend
strongly to specific combinations of features. In the same line of argument,
within-compound associations were proposed to be important in conditioning
experiments to simplify the task (cf. McLaren, Kaye & Mackintosh, 1989).
Next Section: A
Prototypical Point of View
