Relative Importance of Geons
and InterrelationsThere appears to be a substantial role for both object
components and their spatial organization
in the recognition of complex objects or shapes (Brown & Dooling, 1993;
Kirkpatrick-Steger, Wasserman, & Biederman, 1998; Wasserman et al.,
1993; Watanabe & Ito, 1991). This dual role is consistent with Recognition-by-components
(Biederman, 1987), a theory of object recognition in humans, and provides
further support for the notion that parallel processing of object attributes
may occur in avian species as well as in human and nonhuman primates (Brannan,
1992; Goodale & Milner, 1992; Grossberg, Mingola, & Ross, 1994;
Livingstone & Hubel, 1988; Ungerleider & Mishkin, 1982; Zeki, 1993). What remains to be determined
is: (1) what is the relative importance of shape and location information
to object recognition? and (2) is there attentional trade-off between the
control by the two forms of information so that if component (or geon)
shape gains greater control, the amount of control exerted by component
location may suffer?
The examination of the relative control by the geons
and their organization required the construction of a special set of pseudo-objects. Each of the
objects to the right contains two geons: a cube and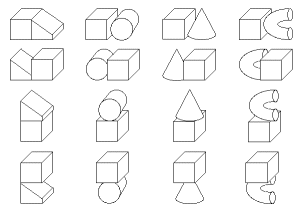 a
variable geon. The variable geon could be one of four shapes: a wedge,
a cylinder, a cone, or a handle. The variable geon could appear in one
of four locations relative to the cube: above, below, left, or right.
a
variable geon. The variable geon could be one of four shapes: a wedge,
a cylinder, a cone, or a handle. The variable geon could appear in one
of four locations relative to the cube: above, below, left, or right.
Kirkpatrick-Steger and Wasserman (1996) trained pigeons to peck at the 16 pseudo-objects.
Each pseudo-object was displayed individually on the viewing screen for 15 s and the first peck after 15 s resulted in
the delivery of a pellet of food. Once the pigeons were pecking at a high rate to
all 16 stimuli (M = 99.7 pecks/min), one of the pseudo-objects was designated as the reinforced stimulus (S+)
and the other 15 were designated as the non-reinforced stimuli (S-). On the S+ trials,
the pigeons would receive food for the first peck following 15 s of stimulus exposure.
On the S- trials, the pseudo-objects were presented for 15 s, but no response was required
and no food was delivered. Four of the pseudo-objects were selected as S+s: handle-below,
wedge-right, cone-above, and cylinder-left. Two birds were assigned at random to received
each of the S+ stimuli.
The differential reinforcement contingency resulted in continued high rates of
pecking to the different S+ drawings and a decrease in response rates to the S-s. The
degree to which pecking to the S- drawings persisted serves as a measure of the degree
of generalization to that S- from the training S+ drawing. Generalization was strongest
to the pseudo-objects that contained either the same variable shape as the S+ or where
the variable shape was in the same location. The next
figure shows the
percentage of total pecks that occurred to the S- stimuli that contained either the correct shape
or the correct location.
Substantial generalization occurred along both the shape and locations dimensions.
different S+ drawings and a decrease in response rates to the S-s. The
degree to which pecking to the S- drawings persisted serves as a measure of the degree
of generalization to that S- from the training S+ drawing. Generalization was strongest
to the pseudo-objects that contained either the same variable shape as the S+ or where
the variable shape was in the same location. The next
figure shows the
percentage of total pecks that occurred to the S- stimuli that contained either the correct shape
or the correct location.
Substantial generalization occurred along both the shape and locations dimensions.
This demonstration of conjoint control by shape and location accords with other demonstrations
of complex stimulus control that have been conducted with a range of experimental procedures and
stimulus dimensions (e.g., Blough, 1993; Chatlosh & Wasserman, 1993; Riley & Roitblat, 1978;
Wasserman, Grosch, & Nevin, 1982). The observed superiority of location information over shape
indicates that spatial interrelations among object components may be more salient than the object
components themselves (although the relative salience may be affected by the particular choice of shapes
and locations that are used in the training set). This is not an unreasonable assertion given that there may be fairly limited
set of primitive components that compose objects (c.f., Biederman, 1987), but a wide range of organizational
rules. As a result, two objects can contain the same components in different interrelations, and be
clearly discriminable from each other (see the cup
vs. pail example).
Kirkpatrick and Wasserman (1996) also observed attentional
trade-off between the shape and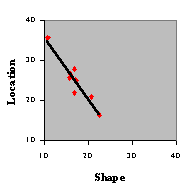 location dimensions (see Chatlosh & Wasserman, 1993 for related evidence). In the bird that demonstrated
greater control by shape, control by location was weaker; in the birds that demonstrated greater
control by location, control by shape was weaker. This is demonstrated in the figure to the right, which
plots the total percentage of pecks to the three S-s that shared the same location vs. the three S-s that shared the same shape as the
S+, for each of the eight birds (this is a replot of the data in the figure above).
The relationship between shape and location was adequately fit by a
decreasing linear function, indicating that there was a negative correlation between attention to shape and
attention to location. The demonstration of attentional trade-off indicates that there may be limits to the
total amount of visual information that can be extracted from a particular set of objects.
location dimensions (see Chatlosh & Wasserman, 1993 for related evidence). In the bird that demonstrated
greater control by shape, control by location was weaker; in the birds that demonstrated greater
control by location, control by shape was weaker. This is demonstrated in the figure to the right, which
plots the total percentage of pecks to the three S-s that shared the same location vs. the three S-s that shared the same shape as the
S+, for each of the eight birds (this is a replot of the data in the figure above).
The relationship between shape and location was adequately fit by a
decreasing linear function, indicating that there was a negative correlation between attention to shape and
attention to location. The demonstration of attentional trade-off indicates that there may be limits to the
total amount of visual information that can be extracted from a particular set of objects.
Next Section: Invariance
