II. Visual Search
When hunting concealed prey, most ground-feeding
birds are engaging in visual search (see
video clip  of
a pigeon searching for grain). It happens that search is widely used
in laboratory studies of visual perception. The laboratory search procedure
requests the participant to find a target embedded in a multi-item display,
which may comprise an array of alphanumeric symbols, forms, colors, or
words. Performance is usually assessed under time pressure; either the
display is flashed only briefly or the participant is instructed to respond
as quickly as possible. An important theme of this chapter concerns the
manner in which such laboratory data and resulting theories apply to natural
foraging.
of
a pigeon searching for grain). It happens that search is widely used
in laboratory studies of visual perception. The laboratory search procedure
requests the participant to find a target embedded in a multi-item display,
which may comprise an array of alphanumeric symbols, forms, colors, or
words. Performance is usually assessed under time pressure; either the
display is flashed only briefly or the participant is instructed to respond
as quickly as possible. An important theme of this chapter concerns the
manner in which such laboratory data and resulting theories apply to natural
foraging.
Some Background
The experimental study of search in humans has a rather different history
than does comparable work in pigeons. Some current research shares common
techniques, however, for which we can thank the computer, especially its
ability to produce quickly an enormous variety of visual displays and to
time events in milliseconds.
Early search studies with humans were motivated by applied considerations
such as the design of radar and other instrumental displays. For example,
Green and Anderson (1956) found that search for a particular numeral
was speeded if the target had a unique color. Over the years,
the search procedure has come to address some deeply theoretical issues
concerning human visual information processing. In summarizing some of
these, it is helpful to return to the distinction between perceptual and
memory variables. Search efficiency is most obviously affected by display
characteristics, such as the density of non-target items. However, search
also depends on the number of potential targets. If, for example, the searcher
is looking for any of several letters, he or she may have to refer back
to memory where representations of the target set are stored. An important
manipulation in search experiments is that of set size, where the set may
refer to either to the number of items in the display or the number of
targets to remembered. These manipulations, along with similarities among
set members, have helped guide modern views about visual-information processing.
Perceptual Search
Human
studies.
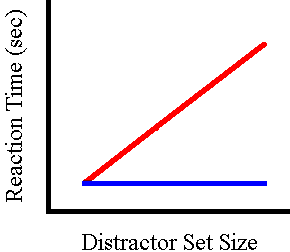
An early and important observation in search studies concerned the
relation between RT and display size. Under some conditions search time
increases with more distractors (the red line in the figure
to the right, which idealizes the distinction
between processes affected by display size and those that are not),
but in others RT seems to change very little with distractor set size (the
blue line). These observations have given rise to a fundamental differentiation
between information-processing styles. When the relation is increasing,
the display items are said to be examined serially or one at time. When
serial processing is literally the case, RT should have a linear relation
to set size, indicating that overall search time is incremented by a fixed
amount as each item is examined. When RT does not change with set size,
processing is said to be parallel; that is, the searcher inspects all of
the items at once; in such cases the search task is easy enough that focused
attention is not required. In fact, truly invariant RTs are rare, but experimental
manipulations distinguish clearly between the two types of function, and
the descriptors are still in use.
Research has not only identified conditions associated with serial
or parallel search but has also led to models detailing the ways in which
humans process complex visual information. For example, manipulation of
similarity among targets and distractors has proven useful. As would be
expected, search is faster when the target is dissimilar to all distractors
in the display, and in cases of sufficient dissimilarity search becomes
almost parallel. A well-known analysis of this phenomenon suggests that
parallel search occurs only when the target possesses a unique feature
that distinguishes it from the other items (Treisman & Gelade, 1980).
According to this analysis, there are a limited number of such features,
which include specific colors and line  orientations. To the observer a
unique feature seems to pop out from the display whether or not it is the
object of attention (see Figure
2). Treisman
and Gelade suggested properties of the human neural system that determine
which visual characteristics constitute features (also see Cook,
2001 for more on this in pigeons).
orientations. To the observer a
unique feature seems to pop out from the display whether or not it is the
object of attention (see Figure
2). Treisman
and Gelade suggested properties of the human neural system that determine
which visual characteristics constitute features (also see Cook,
2001 for more on this in pigeons).
A number of models have attempted to relate such perceptual effects
to stages of information processing. For example, Hoffman (1979) suggested
that easy target-distractor discriminations are analyzed at an early level
that deals with information in parallel. For difficult discriminations
further levels of processing are required, and these are performed in serial.
A modification of Hoffman’s model views the two processes as more closely
blended in time (Wolfe, Cave, & Franzel, 1989).
Avian Studies.
Search in birds as well as humans depends perceptual factors. In foraging
situations similarity between prey and background is a natural condition,
because such crypsis is beneficial to the prey. Studies that address foraging
strategies are most likely to assess consummatory behavior, using observational
techniques and prey appropriate to the species under investigation. For
example, Dawkins (1971) observed chick feeding behavior when dyed grains
were scattered on surfaces that were similar or different in color. Her
findings showed that the birds initially consumed the concealed grains
more slowly, although their speed increased with practice. Gendron (1986)
reported a similar outcome in observations of quail selecting food pellets.
Of course, these results are hardly surprising, and it should be noted
that such comparisons were secondary to the main purpose of the cited research.
The section on search image return
to this and related work
Other research approaches substitute simple pecking for the full consummatory
response. For example,  experiments with jays required the birds to peck
at photographic displays when the image included a moth; the moth, whenpresent,
could appear against a similar or dissimilar natural background (Pietrewicz
& Kamil, 1979). In our laboratory, pigeons peck at computerized displays
made up of artificial symbols (See Figure
1) .
experiments with jays required the birds to peck
at photographic displays when the image included a moth; the moth, whenpresent,
could appear against a similar or dissimilar natural background (Pietrewicz
& Kamil, 1979). In our laboratory, pigeons peck at computerized displays
made up of artificial symbols (See Figure
1) .
As in the human studies noted earlier, we have explored target concealment
by varying the number of distractors and their similarity to the
targets. Figure
4 illustrates some simple display-size
and similarity effects obtained in a pigeon study (Blough, 1992). The
distractors were alphanumeric symbols. In a conspicuous or low-similarity
condition the target was a filled heart. In intermediate and high-similarity
conditions targets were a '/' and a 'B' respectively. As shown below, search
for the filled form was faster overall, indicating that this target was
the most distinctive. Further, the mode of search appeared to be parallel,
contrasting with the seemingly serial style associated with the less distinctive
targets.
display-size
and similarity effects obtained in a pigeon study (Blough, 1992). The
distractors were alphanumeric symbols. In a conspicuous or low-similarity
condition the target was a filled heart. In intermediate and high-similarity
conditions targets were a '/' and a 'B' respectively. As shown below, search
for the filled form was faster overall, indicating that this target was
the most distinctive. Further, the mode of search appeared to be parallel,
contrasting with the seemingly serial style associated with the less distinctive
targets.
Other perceptual factors include the way in which display items are
grouped; for example, detection seems faster if distractors are alike (Blough
& Blough, 1990). Natural crypsis is enhanced by the clutter characteristic
within foraging patches.
It is interesting to ask whether there are certain natural "features"
that command the pigeon's attention. Treisman and Gormican (1988) placed
forms distinguished by a line or a gap in that category; that is, search
appeared parallel for such forms, but was "serial" when the form
was distinguished instead by the absence of a line or a gap. and this finding
was replicated with humans in our search paradigm by Allan and Blough (1989).
However, when pigeons performed a comparable search task, there was no
evidence for such special properties. Of course, these findings do not
rule out the possibility of special "pigeon features."
Memory Search
Human Studies.
Memory assumes a special role in visual search when the observer’s
task is to locate any of several targets. In such cases he or she will
presumably refer to a list stored in memory, and the task can be labeled
memory search. Of course, memory search occurs in everyday circumstances
as well. A familiar example occurs when one tries to recall the name associated
with an acquaintance’s face.
Again research has distinguished between parallel and serial processes,
marked by set-size effects. In a search paradigm, Schneider & Shiffrin
(1977) varied the strength of the memory demand. In advance of each trial
participants viewed a list specifying potential target items; the
size of this memory set was variable. Following a subsequent test display
the viewer indicated whether or not one of those targets was present in
the display. In a relatively easy "consistent-mapping" task, members of
target set were the same throughout the session. In a more difficult "varied-mapping
task," symbols that had been targets on one trial could appear as distractors
on another. Search appeared to be parallel in the consistent condition;
that is, the practiced observer's speed did not depend on the number of
items in the list. In contrast, search time increased with the number of
items in the variable condition.
One way of understanding the difference between consistent and varied
memory sets invokes the distinction between working and reference memory.
Working memory has a limited capacity and is transient; it holds new information
that has just been supplied or old information recently refreshed. Reference
memory is relatively permanent, representing a vast amount of previously
acquired knowledge. Schneider and Shiffrin’s (1977) varied-set task taps
working memory, because the searcher must address new information at the
start of each trial. Information required for the consistent-mapping task
could remain in reference memory, because the distinction between
targets and distractors was constant throughout a session,. The differences
in set-size functions are understandable if we assume that working
memory is addressed by a serial process while reference memory is accessed
by some form of parallel process (e.g. Logan & Stadler,1991.).
Closer scrutiny of RTs in a fixed-set task shows an overall reduction
with practice (Shiffrin & Schneider, 1977). This change may be attributable
to a gradual disengagement from working memory as the searcher learns the
distinction between targets and distractors. If this interpretation, as
well as those suggested above, are correct, the set-size effect should
be present early-on and should diminish with experience as long as a consistent-mapping
task is used. This changeover is difficult to capture in humans because
of the speed with which it seems to occur.
Avian
Studies. Because they consume varying grain types, foraging
pigeons may also address a memory list of acceptable targets. It is also
reasonable to distinguish between lists that reside in working and reference
memories. Avian research that has operationalized this distinction has
helped us understand set-size effects seen in the pigeon search task.
An early analysis (Honig, 1978) recalls the Shiffrin
and Schneider (1977) distinction between fixed and variable memory sets.
In particular, Honig distinguished between aspects of a task that varied
between trials  and those that remained fixed over a session. The matching-to-sample
experiment provides a good illustration (see demo
from Grant
& Kelly, 2001).
The sample changes from trial to trial, so its current value is variable
and would be held in working memory. However, the relation between a given
sample value and the associated response remains constant, and that information
could remain in reference memory.
and those that remained fixed over a session. The matching-to-sample
experiment provides a good illustration (see demo
from Grant
& Kelly, 2001).
The sample changes from trial to trial, so its current value is variable
and would be held in working memory. However, the relation between a given
sample value and the associated response remains constant, and that information
could remain in reference memory.
Several approaches have considered the capacity of pigeon working memory
and the nature of information that is retained when the load is high. Chase
(1983) showed, for example, that pigeons can correctly identify up to 3
items just seen; Shimp (1976) and Terrace
(2001) have described
the correspondence between what is remembered and the order in which information
is presented. Such work helps to define the contents of working memory;
the search procedure should allow us to address the manner in which they
are addressed. In a "conditional" search procedure we attempted to specify
the effects of set size on search RT (Blough & Blough, 1990). Here
letter stimuli could be either targets or distractors; pre-trial cues informed
the bird which letters would be targets in the forthcoming display. In
some conditions the cue was mapped to a single letter; in others it was mapped to a set of several targets, only one of which appeared. The
data suggest a serial mode; that is, RT increased with the number
of targets cued, but the effects are small and need replication for a clear
conclusion.
As in humans, pigeon long-term memory has a relatively large capacity.
Tests of picture recognition, for example, indicate that birds can categorize
large sets of photographic slides according to whether or not they have
been seen previously (Vaughan & Greene, 1983).
It is relatively easy to conduct a fixed-set memory search task with
pigeons, and we have explored this matter. In the display-search task illustrated
earlier
pigeons are rewarded for pecking at any of several target letters, which
are drawn from a fixed set. Only one of those targets appears in any given
display, and the number of letters in a set varies in different phases
of a study. If pigeons as well as humans address reference memory in parallel,
RT should not depend on set size. An early study (P. Blough, 1984) provided
seemingly inconsistent results. Experiment 1 revealed an RT elevation associated
with the larger of two set sizes. However, the effect was not replicated
in Experiment 2, conducted with the same birds and some of the same target
letters. We speculated that practice improved the efficiency with which
the birds accessed a multiple-item list.
In subsequent work, Vreven and Blough (1998, Experiment 2) considered
set-size and practice effects more fully. Birds learned to search for each
of four targets, but in sessions that presented only a single letter or, in a  mixed
condition, any of four letters selected randomly. Test
sessions then presented either single letters as well or, in a mixed condition
where all four letters were presentedrandomly
within a session. The data reveal a sizeable set-size effect during
initial test sessions, but RTs for the two set sizes eventually converged
(See Figure 5).
This outcome is consistent with the notion that practice brings about changes
in the way multiple memory items are processed and that a switch from working
to reference memory may be involved.
mixed
condition, any of four letters selected randomly. Test
sessions then presented either single letters as well or, in a mixed condition
where all four letters were presentedrandomly
within a session. The data reveal a sizeable set-size effect during
initial test sessions, but RTs for the two set sizes eventually converged
(See Figure 5).
This outcome is consistent with the notion that practice brings about changes
in the way multiple memory items are processed and that a switch from working
to reference memory may be involved.
It is logical to think that memory search would proceed in a different
manner if some of the targets in a set were similar to each other. For
example, a search task might employ targets that shared features with each
other, but not with any of the distractors. In that case, the common feature,
if identified, would constitute a single target, and the set would have
a size of one. Our experiments were designed to avoid this situation, but
it reasonably could occur in a foraging patch. Two search-image studies,
to be described in the next section, illustrate cases in which birds seemed
to generalize between targets.
Next section: Attention and the
Search Image