III. Attention and The Search
Image
In a natural foraging situation, a bird's most effective strategy maximizes
its food intake while minimizing energy expenditure. Of course there are
various ways to achieve this balance, but, for walking birds, costs are
reduced when prey appearing within the current field of view are detected
quickly. This section is devoted to a widely-studied perceptual
mechanism, attention, that can speed detection of a particular target,
but, at the same time, reduces the chances that alternative targets will
be noticed.
A series of field studies by L. Tinbergen (1960) inspired much of the
avian research and theory that will be summarized here. At a site within
a Dutch forest Tinbergen monitored great tits (a relative of the chickadee)
as they returned from foraging trips to their nest boxes. Through analysis
of the remains of a captured prey, he related characteristics of recovered
prey species to days and times of year. In this setting the densities of  individual species in the forest varied according to the season, so it
isn't surprising that the numbers of captured prey reflected its natural
density for the corresponding time period. Rather, the outcome of
interest was the disproportionately large numbers of
captures relative to
an abundant prey's actual density. This tendency to overselect the
more common species suggests that the tits’ perceptual systems were biased
toward unique features characterizing such prey. Figure
6 illustrates this sort of bias.
individual species in the forest varied according to the season, so it
isn't surprising that the numbers of captured prey reflected its natural
density for the corresponding time period. Rather, the outcome of
interest was the disproportionately large numbers of
captures relative to
an abundant prey's actual density. This tendency to overselect the
more common species suggests that the tits’ perceptual systems were biased
toward unique features characterizing such prey. Figure
6 illustrates this sort of bias.
Tinbergen used the term, "specific searching image, (p.332) to account
for his observations. The "image" was induced by repeated encounters with
a given prey, and its' presence improved the tit's ability to notice a
similar insect on a subsequent flight. Tinbergen's discussion invoked concepts
from both the learning and perceptual traditions. The fact that overselection
appeared to grow with repetitions of a given prey type suggested learning
or conditioning. Yet the searching image is described as a "selective sieving
operation on the visual stimuli...." (p 332), implying the filter-like
process that characterizes attention.
Among the many laboratory studies that have examined Tinbergen's hypothesis
is work by Pietrewicz and Kamil (1979) and by Bond (1983). Pietrewicz
and Kamil required blue jays to respond differentially to photographs of
trees depending on whether or not the picture included a concealed moth
resting on the bark. There were two moth species and each rested on bark
having a similar appearance to its own. To mimic repeated encounters, short
trial sequences presented only a single species (a "run"). Control sequences
presented the species in random order. This work conceptually replicated
Tinbergen's observations, because the jays were more accurate during the
runs. The work also helped rule out certain alternative accounts of overselection;
for example, the effect could not be attributable simply to practice in
handling a particular prey type or to improved travel efficiency.
Bond's (1983) work addressed the distinction
between attentional and conditioning accounts of overselection. His approach
emphasized the matter of crypsis. If a search image is attributable to
improved attentional focus, overselection should be more pronounced under
cryptic conditions; focused attention would not be required for conspicuous
prey. If learning over repeated encounters improves detection, however,
there should be no effect of crypsis; if anything, performance should improve
more slowly over encounters with poorly-discriminable stimuli. Bond's experiments
provided pigeons with opportunities to select grains from food trays containing
two types in varying proportions. In one condition a variegated substrate
resembled the grains and, in another, the background was relatively homogenous
and distinctive. In the latter or conspicuous condition the birds tended
to match their choices to the density of the grain type; that is their
choices resembled the linear function shown in figure above. In the cryptic
condition, however, they overselected a given type when it was moderately
abundant; that is, their choices resembled the S-shaped functions shown
in Figure
6. These data, then, were most
consistent with an attentional than with a reinforcement account of search
images.
Human Attention
As suggested above, the search image might be best understood as an
attentional process. This section highlights certain ideas found in this
vast literature. The usual definitions describe attention as a capacity-limited
resource that, when focused on a perceptual or memory task, improves
performance. Because of the capacity limitation, however, unattended tasks
are performed less well than would be the case if attention were not engaged
at all. In other words, there are costs as well as benefits to focused
attention.
A second important point is the distinction between situations that
do and do not benefit from attentional focus. Operational distinctions
between these are often based on the criteria for serial and parallel search,
summarized earlier. The idea of focused attention is implicit in serial
processing; that is, attention is directed toward one memory or display
item at a time. In parallel search all members of either set are processed
at once without focal attention. In the case of perceptual tasks, as noted
above, good discriminability favors parallel search. In the case of memory
tasks, a condition favoring parallel search is an fixed or unchanging memory
set.
Studies of the human attentional process may use various techniques
to engage focal attention; typically, the task is made difficult, for example,
by concealing targets or flashing displays very briefly. One illustration
comes from the psychophysical tradition, which requires an observer to
distinguish among very similar stimuli along a specified dimension. In
a study of contrast sensitivity, Davis, Kramer, and Graham (1983) required
observers to discriminate gratings of varying contrasts and spatial frequencies.
In one condition the same spatial frequency was repeated over a series
of trials, while, in another, frequencies were intermixed. This method
resembles Pietrewicz and Kamil's (1979) runs-mixed method, and the results
were also similar; that is, accuracy was higher when trial types
were repeated.
In studies of human perception, Posner and Snyder (1975) required participants
to report the identity of displayed letters. Preceding each display
a symbolic cue appeared. In some conditions this signal predicted the identity
of the forthcoming target letter on a large percentage of the trials; in
others the signal was ambiguous; that is, it was valid on just half the
trials. Performance was faster when the cues were valid than when they
were ambiguous. Further, miscues (trials when the "valid cue" did not predict
accurately) were associated with RTs still higher than those associated
with the ambiguous condition. Posner and Snyder interpreted this
work in terms of attentional primes: The miscues as well as the valid cues
(primes) directed attention to a particular form, while the ambiguous cues
presumably did not direct attention at all. The comparison between
ambiguous cues and miscues is especially important, because it demonstrated
a cost of misdirected attention.
Only some of the reports mentioned so far emphasized "attentional" processes.
Davis, Kramer and Graham (1983) attributed the poorer performance associated
with mixed stimulus sequences to "uncertainty," while Pietrewicz
and Kamil (1978) attributed their findings to the absence of a searching
image . It seems reasonable to ask, however, whether the improved performances
in all these cases result from a common process. One could argue, for example,
that effects produced by trial repetition are different from those yielded
by the symbolic cues employed in some parts of Posner and Snyder’s work.
For example, stimulus repetition could exert a "bottom-up" influence; that
is, a persisting stimulus trace may facilitate the detection of a matching
target should it appear within a short time. Symbolic cues, which predict
but do not match the forthcoming target, would engage a "top-down" process,
since their significance would require more interpretation.
The next section emphasizes our studies of search images in pigeons.
It will consider the relationship between repetition-induced "search images"
and symbolic attentional primes, as well as the matter of bottom-up
influences and the role of learning.
Avian Attention
Our studies of attention during visual search were motivated initially
by the work, mentioned above, of Tinbergen (1960), Pietrewicz & Kamil
(1979) and Bond (1983). Those studies, guided by naturalistic considerations,
measured probability of prey detection or capture. In our laboratory
we stressed search RTs, because of their exceptional sensitivity to perceptual
and memory variables. Much of this work was driven by the hypothesis that
a “search image” is a primed representation that facilitates detection
of the represented item in a noisy background
The role of
similarity. The
literature on search images emphasizes a “cryptic requirement;” that is,
prey concealment favors the facilitory effect of abundancy. Bond’s studies
and others (e.g. Dawkins, 1971; Bond & Kamil, 1999) have provided support
for such a requirement, and it is also consistent with human data suggesting
that attention is mostly likely to be engaged when perceptual demands are
high.
Some of our experiments have also considered
similarity by manipulating the relation between target and background letters
and have raised questions about the generality of the cryptic requirement.
For example, priming both by runs and discrete cues was effective over
a wide range of display sizes, and there was no evidence that the size
of the effect interacted with the number of display items (P. Blough,
1989, 1991). In other work repetition priming was seen in the case of an
especially salient target, a filled heart among alphanumeric distractors
(P. Blough, 1992). We can only speculate about the differences between
our data and others; it is possible, for example, that our RT measure is
especially sensitive to attentional effects and reveals them at lower levels
of similarity than other measures. Indeed, the experiments just cited showed
“ceiling effects” on accuracy; that is, the birds detected the target on
most trials, but they did so with varying speeds.
The role of bottom-up factors. Some
recent studies have considered the contribution to priming of brief memory
traces carried over from trial to trial. For example, within same-target
sequences, a trace of the just-seen stimulus could enhance detection of
the repeated target, especially if trials were closely spaced. Bond’s analysis
(1983) implicated this bottom-up account; according to his model, capture
of a given prey triggered a switch to a specific attentional mode that
lasted only briefly. More recently, Plaistead (1997) found that short intertrial
intervals facilitated search for simulated prey displayed on
a video screen and that target repetitions enhanced this effect. By an
argument somewhat different from Bond’s, she also concluded that transient
memory traces contributed to the runs effect.
In contrast to the analyses of Bond and Plaistead, there are reasons
to believe that transient stimulus traces do not account for all cases
of attentional priming. For example, other studies of sequential priming
(P. Blough, 1991; Bond and Kamil, 1999) did not reveal intertrial interval
effects; these studies, like Plaistead’s (1997), manipulated such intervals
over a range of a few seconds. In field studies such as Tinbergen’s (1960),
substantial amounts of time elapsed between foraging trips; indeed, as
Bond and Kamil point out, search images would be beneficial only if they
lasted over a realistic travel time.
On the other hand, facilitation by stimulus traces could be evident
when a variety of targets are encountered at random and sequences within
which a given target happens to be repeated are compared with those within
which different targets appear. Compare, for example, the following 5-trial
sequences:
|
Sequence 1 = B-D-A-C-B
Sequence 2 = D-A-C-B-B
|
Especially if trials are closely spaced, a stimulus trace could facilitate
the detection of the final B in the second sequence.We have explored this
possibility by analyzing data collected in the laboratory search task.
In one procedure we simply varied the probabilities with which members
of a 4-target set appeared; that is, during each session, the designated
frequent target appeared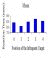 with a probability of 0.9, and
an alternative target appeared with a probability of 0.3 (P. Blough, 1996).
This method offered opportunities to compare RTs for targets that had and
had not been seen on just-preceding trials. Figure
7 shows the outcome of one such analysis, which provided no evidence
for speeded detection following target repetition.
with a probability of 0.9, and
an alternative target appeared with a probability of 0.3 (P. Blough, 1996).
This method offered opportunities to compare RTs for targets that had and
had not been seen on just-preceding trials. Figure
7 shows the outcome of one such analysis, which provided no evidence
for speeded detection following target repetition.
In a more systematic approach, we hypothesized that repetition
facilitated search only when it provided information about target identity
(Blough & Lacourse, 1994). For example, in the runs-mixed comparison,
a sequence of two or more target repetitions signals that a run is probably
in effect and thus provides information about forthcoming target identity.
Facilitation in this case could result from a priming mechanism similar
to that seen in discrete predictive cues. Our experiment compared groups
of three trials in which the same target appeared. In the predictive condition
targets were regularly repeated in sets of three and then changed; in a
random control we selected targets at random, but pulled out sets of 3
repetitions that occurred by chance within the random design. In both conditions,
RTs declined over the 3-trial series, but the decline was greater in the
predictive condition. Although the difference was not large, these data,
together with the analyses mentioned above, suggest that the informative
value of runs or differential target probability enhances their facilitory
effects. Such an account implies a top-down mechanism, because to
generate a prediction, the priming event must activate memory.
The role of
learning. Tinbergen
supposed that “a specific searching image is assimilated when the birds
have had a number of chance contacts with the species in question.” (Tinbergen,
1960, p. 316). While he and others speculated that learning was involved,
the nature of the exposure effect is still not clear. While we have proposed
an attentional mechanism, some form of conditioning could also contribute.
When the runs procedure is used to induce a search image (e.g., Pietrewicz
and Kamil, 1979) it is possible that differential reinforcement is responsible
for the improved performance; that is, short-term practice effects could
occur within a same-target sequence. Pietrewicz and Kamil minimized, but
did not eliminate, this possibility by balancing, within a session, the
number of trials associated with each member of the target set. A
practice effect is not inconsistent with Tinbergen’s (1960) views, but,
in humans, attentional switches are induced by very brief events, such
as the discrete cues used by Posner & Snyder (1975). We considered
the effects of such cues with pigeon in a design that presented briefly
a symbolic form just before onset of the search display. A separate distinctive
form predicted the identity of each of two targets on 75% of the trials;
in those cases the forms were were valid cues, but on the remaining 25%
of the trials they were miscues. A third form predicted target identity
on 50% of the trials and thus was an ambiguous cue. Compared to the ambiguous
condition, the valid cues increased detection speed much as runs do.(P.
Blough, 1989, 1991). Although the cue-target association is acquired through
practice, its facilitating effects on search cannot be attributed to differential
reinforcement. In some of this work ( P. Blough, 1991) we attempted to
specify the role of attention by assessing costs and benefits of the two
procedures. In the symbolic cue method, miscues produced higher RTs not
only than did valid cues, but also when compared to ambiguous cues.
An analysis applied to the runs procedure followed a related logic: To
establish costs, a new target following a run (“miscues”) should
have produced higher RTs than did “ambiguous” targets (those just following
a mixed sequence). The effects followed the predicted pattern, but the
differences were not significant. Thus these experiments established the
attentional change induced by discrete cues, but they did not fully clarify
the nature of the runs effect.
In order to explore that mechanism further, we turned to a related procedure.
In nature, birds will encounter the abundant prey more often, but in varying
rather than regular sequence lengths. To mimic this arrangement our design
varied target probability (P. Blough, 1996). Figure
8 illustrates an outcome of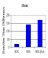 such
an experiment. The lower RT associated with the more probable target provides
good evidence for improved detection as a result of repeated encounters,
and an incentive explanation might seem reasonable; that is, the more probable
target is associated with more frequent reinforcement during a session.
However, other data from our laboratory (D. Blough, 1992) dissociated the
effects on RT of these two reinforcement variables. In a design that varied
them independently, RT declined with increasing reinforcement probability,
but not its frequency. In the present experiment reinforcement probability
was constant over target probability conditions and could not have been
responsible for the differential RTs.
such
an experiment. The lower RT associated with the more probable target provides
good evidence for improved detection as a result of repeated encounters,
and an incentive explanation might seem reasonable; that is, the more probable
target is associated with more frequent reinforcement during a session.
However, other data from our laboratory (D. Blough, 1992) dissociated the
effects on RT of these two reinforcement variables. In a design that varied
them independently, RT declined with increasing reinforcement probability,
but not its frequency. In the present experiment reinforcement probability
was constant over target probability conditions and could not have been
responsible for the differential RTs.
Another aspect of this design permitted a further test of a reinforcement
account. Within a session, the identity of the most likely target was constant,
but it changed between sessions according to a random program. On most
days, the birds were rewarded occasionally for a correct response. However,
occasional test sessions omitted all reinforcement. As shown in the figure
above, the probability effect persisted during these tests. (P. Blough,
1996).
Recall that Bond (1983) also provided a compelling argument against
a reinforcement account of the search-image effect (click
here to review Bond's argument). Overall, it appears that detection improves with exposure, but not because
of a reinforcement-based strengthening of the target-response association.
The role of exposure. Much
of our work, as well as that from other laboratories, has suggested that
advance information about target identity acts a priming event; such information
can be provided by valid symbolic cues as well as uninterrupted runs. In
recent unpublished analyses we have considered evidence suggesting that
exposure alone facilitates detection.
These studies varied target-probability and explored the course of RT
over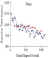 lengthy sessions after the manner reported in P. Blough, 1996. In this
work there were only two targets whose relative probabilities varied between
sessions. Figure 9
illustrates such functions in conditions that set probabilities at 0.7
and 0.5. As shown, RTs in both conditions dropped over the session in a
negatively-accelerated fashion and additional analysis revealed similar
drops over a range of target probabilities. It is clear that a within-session
increase in response speed is not confined to targets having a high probability
or to the presence of reinforcement.
lengthy sessions after the manner reported in P. Blough, 1996. In this
work there were only two targets whose relative probabilities varied between
sessions. Figure 9
illustrates such functions in conditions that set probabilities at 0.7
and 0.5. As shown, RTs in both conditions dropped over the session in a
negatively-accelerated fashion and additional analysis revealed similar
drops over a range of target probabilities. It is clear that a within-session
increase in response speed is not confined to targets having a high probability
or to the presence of reinforcement.
Finally, Figure
10 summarizes RTs for high and low probability conditions, showing
elevations only at  relatively lowfrequencies
of occurrence. A simple account of effects may be drawn from Figure 9.
Because those functions are negatively accelerated, RTs tend to converge
when there has been a sufficient number of exposures. When exposures are
relatively few, as in the case in the low-probability conditions, an RT
differential persists.
relatively lowfrequencies
of occurrence. A simple account of effects may be drawn from Figure 9.
Because those functions are negatively accelerated, RTs tend to converge
when there has been a sufficient number of exposures. When exposures are
relatively few, as in the case in the low-probability conditions, an RT
differential persists.
The data are consistent with Logan’s (1988) observations that RTs associated
with a variety of human tasks decline with exposure in an exponential manner.
To account for such effects, Logan (1988) invoked a gradual switch from
a slow to a fast processing style. The present data do not address the
adequacy of that explanation. However, as noted above, our previous work
does rule out an reinforcement account as well as one that assigns a facilitating
effect to brief stimulus traces.
The role of extended experience with
multiple search targets. In an earlier discussion of avian memory
search, a rather different effect of experience was noted. It was seen
that the single-target advantage diminished over the course of extended
practice (Vreven & Blough, 1998). A plausible explanation suggests
that attentional demands decline with experience in a given task. In other
words, the experienced searcher may adopt a fast “automatic” or "parallel"
processing style, perhaps as the result of altered processing strategies
attributed to Logan (1988). Recall from our discussion of human
memory search that such a strategy would be limited to fixed memory sets
such as those used in our pigeon experiments.Some of our evidence suggests that parallel search is less successful
with larger memory sets. In our study of memory search (Vreven &
Blough, 1998), practice effects did not emerge for sets of 8 to 12 items.
There may be capacity limitations to the parallel process that searches
automatically.
In summary, we believe that the search image, originally postulated
on the basis of naturalistic observations, may be an instance of improved
attentional focus achieved through exposure to a particular prey type.
However, the processes that modulate this change are not simple any more
than is the world in which most birds conduct their foraging activities.
Natural conditions that favor search image formation are most likely those
that present a variety of prey types that vary in density and whose identities
change over time. It is likely that nature also provides foraging patches
so simple and familiar that selective attention is not helpful. As in other
areas of avian cognition, our understanding will advance most effectively
through collaboration among those scientists who observe birds in nature,
those who simulate natural conditions in their laboratories, and those
who attempt more precise control through artificial means.
Next section: References
