|
|
|
Contrasting views of avian attention cause a tension in
both laboratory science and popular culture. For much of this
century, laboratory science has usually interpreted a bird’s
"mental life" in terms of impersonal, objective,
mechanical "value-free" processes, such as conditioning.
At the same time, however, a minority view has held that a bird’s
mental life bears striking similarities to that of a human, so that
empathic understanding of a bird’s mental life might to some
degree be conceptually meaningful and empirically possible.
Similarly, popular culture seems inclined sometimes to adopt an
impersonal approach to birds and at other times to interpret a bird’s
mental life, especially that of bird pets, in more mentalistic or
subjective and "value-laden" terms, including empathic
understanding, attachment and bonding.
This tension motivates many basic questions. What is the avian
mind? Are there profound similarities between human and avian minds
in terms of objective, impersonal issues? If so, do these
similarities imply there are also similarities in terms of
subjective, personal feelings?
Contemporary laboratory research on avian cognition addresses
these questions. This research is identifying new similarities
between avian and human cognition. As examples of this research, we
describe three similarities between avian and human attention,
specifically in spatial attention, local/global attention, and
selective attention. We explore how similarities such as these
between avian and human attention exacerbate growing tensions
between value-free and value-laden views on the relation between
animal and human intelligence: They amplify unresolved
contradictions in science and in Western culture involving the
deepest ethical, economic, religious, and political assumptions.
These similarities, contradictions, and tensions are addressed in
a class called "The Animal Mind in Nature, in the Laboratory,
and in Human Culture," offered in the Psychology Department at
the University of Utah. Students in this course study the science of
comparative cognition as well as volunteer service at the Hogle Zoo,
Tracy Aviary, Salt Lake Dog Training Club, State Division of
Wildlife Resources, Hawk Watch International, the Humane Society, or
Salt Lake County Animal Services. The course examines ties between
laboratory science on nonhuman animal cognition and the larger
cultural context. We hope the class promotes greater public
understanding of the science of nonhuman animal cognition, and at
the same time, we expect that an examination of the influence of
cultural beliefs on the development of science ultimately will
illuminate theoretical assumptions not yet systematically evaluated
or even yet identified by contemporary research on comparative
cognition |

I. Introduction
Contrasting views of avian attention cause tension in both
laboratory science and popular culture. For much of this century,
laboratory science has usually interpreted a bird’s "mental
life" in terms of impersonal, objective, mechanical
"value-free" processes, such as conditioning. The chapter
in this volume by Sutton and Roberts
(2001) contains a clear historical review of this approach. At the same time,
however, a minority view has held that a bird’s mental life bears
striking similarities to that of a human, so that empathic
understanding of a bird’s mental life might to some degree be
conceptually meaningful and empirically possible. Similarly, popular
culture seems inclined sometimes to adopt an impersonal approach to
birds and at other times to interpret a bird’s mental life,
especially that of bird pets, in more mentalistic or subjective and
"value-laden" terms, including empathic understanding,
attachment and bonding.
This tension motivates asking, What is the avian mind? What
objective similarities are there between human and avian minds? Are
there corresponding subjective similarities? Finally, is it possible
that there is an interaction between the value-free, scientific
approach and the value-laden popular approach? If so, can we make
any predictions about how these two different approaches may affect
each other in the near future?
Research on avian cognition is identifying new similarities
between avian and human cognition, three of which we describe;
spatial attention, local/global attention, and selective attention.
We explore how similarities such as these affect the tension between
value-free and value-laden views on the relation between animal and
human intelligence and we explore how these similarities amplify
unresolved contradictions that have deep ethical, economic,
religious, legal, and political ramifications.
These similarities, contradictions, and tensions are addressed in
a class, "The Animal Mind in Nature, in the Laboratory, and in
Human Culture," that has been offered in the Psychology
Department at the University of Utah. Students study the science of
comparative cognition and volunteer service at the Hogle Zoo, Tracy
Aviary, Salt Lake Dog Training Club, State Division of Wildlife
Resources, Hawk Watch International, the Humane Society, or Salt
Lake County Animal Services. We hope the class promotes public
understanding of the science of comparative cognition, and at the
same time, we expect that an examination of the influence of
cultural beliefs on the development of science ultimately will
illuminate theoretical assumptions not yet systematically evaluated
or even yet identified by contemporary research on comparative
cognition.
 We
wish to alert the reader at the outset that this chapter can be
viewed rather like a verbal example of a classic reversible figure,
such as Figure 1 (left). A reader can see two very different
chapters depending on the reader’s primary interests. One way of
looking at the chapter reveals material on avian visual attention
and the relation between scientific literatures on avian and human
visual attention. A second way reveals material on the avian mind
from the perspective of human culture. A reader chiefly interested
in the first way of seeing the chapter should concentrate on section II. A reader interested in the second perspective
should concentrate on section III. We feel, however, that
neither perspective alone captures our chief point, which is that
science and culture interact, with each importantly shaping the
other. We hope, therefore, that the reader will try to transcend
seeing our chapter as either on the science of avian attention or on
how our culture views the avian mind, or even on both of these
themes viewed separately, and will try instead to think about how
science and culture mutually affect each other.
II. Attention in Science and Culture
Our aim is to present a few examples of research on avian visual
attention, show three ways avian attention resembles human
attention, and to highlight some dramatic and increasingly
problematic issues that similarity exacerbates. We aim to make
explicit how polarized are our culture’s views on the relation
between humans and nonhuman animals, with some individuals,
including some scientists in the context of their professional
lives, having a dispassionate and impersonal point of view on
nonhuman animals, and with other individuals (including some
scientists outside of their professional lives) having sharply
contrasting passionate and highly personal views, especially of
pets. Nonhuman animals, especially pets, seem more "human"
in the latter case than in the former. This polarization sets the
stage, we suspect, for unhealthful contests between
"objective" and "empathic" views of nonhuman
animals. Tension between these views is becoming intolerable for
some members of the public, who are increasingly resorting to
demonstrations, protests, and even civil disobedience of varying
levels of violence. We hope that openly acknowledging the
polarization will facilitate constructive discussion of its origins,
of its scientific and political implications, and possible means of
reconciliation, or at least, constructive management of the
polarization.
Seldom are there polar opposites that could benefit more from a
calm consideration of each other’s methods and goals than in the
case of personal and impersonal views of nonhuman animals. In the
context of today’s polarized positions on the relation between
human and nonhuman minds, even such a moderate position as the one
we just articulated in the previous sentence can seem radical. We
therefore hasten to correct two possible misinterpretations of our
goal. We suspect the first misinterpretation might arise in the mind
of a behavioral scientist: By suggesting that individuals on each
side of this polarized dimension consider the wisdom represented at
the other pole, we do not mean to advocate the replacement of avian
cognition as a science by a kind of folk ethology. We do not mean to
suggest that experimental research is irrelevant to progress on
understanding how human values shape attitudes about nonhuman
animals. Witness the analogous case in which a better scientific
understanding of the role of DDT on the environment contributed to
shaping the public’s values on environmental issues. Nor do we
advocate a return to the use only of anecdotes, intuition, or the
application of unexamined traditional beliefs to legitimize either
any position on the nature of nonhuman animals or on appropriate
human values associated with nonhuman animals. We advocate only that
it would be useful to ask how the impersonal, logical rigor of
science and deeply personal human values might mutually inform each
other in the specific context of the role of nonhuman animals in
human society.
We suspect the second misinterpretation might arise in the mind
of a more general reader. Such a reader might wish for a deeper
commitment to a more humane, subjective, personal, emotional, and
less rigorously scientific perspective than the one we adopt. We
hope such a reader will at least be gladdened by our attempting to
raise the issue of human values in a scientific context and will be
patient with our aim of only hoping to identify questions, not to
answer them.
Consider a set of analogous polarized issues familiar to everyone
affected by how cities have developed in the 20th
century. Personal and impersonal approaches are frequently in
conflict when the future development of cities is discussed, but
increasingly, it is seen that they do not need to be. Cities can be
viewed as vast, impersonal machines to be understood in terms of
numerically described traffic patterns, population densities,
reaction times by fire departments to emergencies, the hydraulics of
the delivery of safe drinking water, structural integrity of tall
buildings, and so on. Alternatively, cities can be seen as human
living places where people experience the deepest human cultural
values by listening to Mozart’s The Magic Flute, enjoying
picnics in public parks, and perhaps even by discussing the nature
of nonhuman animals. While these two visions of cities are often in
conflict, just as are the two visions of nonhuman animals, we see no
necessary conflict between them. To design cities on a human scale,
to improve the quality of life in cities, to reduce environmental
pollution, and so on, in a word, to acknowledge human values in the
context of city life, does not require the abandonment of, and in
fact, we believe requires the use of, meteorological science,
structural engineering, computer simulation of traffic flow, the
delivery of health care based on biological science, and the full
range of science-based technology. Similarly, we believe that a
fuller understanding of the human-ness, or lack thereof, of nonhuman
animals will require attention to both science and human values. We
expect it will not go unnoticed that the question of what a nonhuman
animal is, as we have presented it, closely resembles many analyses
in the "culture wars" fought over differences between
"scientific" and "cultural" approaches (Hacking,
1999). We believe impersonal and personal accounts of the role of
nonhuman animals in human society have been in conflict for a
sufficiently long time that it is time to try something else. We are
optimistic that this particular war can and should be
"fought" at the level of tolerant discourse in which
participants on both sides struggle to understand and respect the
opposing side, not to crush it.
Our more specific goals include the following. We propose to
review three ways in which attention has been studied in human
cognitive psychology. We then show how these same well-established
attentional processes can also be studied scientifically in avians.
We note how this research on both human and avian attention is
typically seen as largely objective and independent of human values.
We note how the very success of this laboratory research, which
makes avians seem more like humans, raises currently unanswered
questions about the assumption that attentional processes in avians
are best viewed solely in entirely objective terms. To emphasize the
current unresolved polarization between objective and empathic views
of nonhuman animals, we list some mutually incompatible implicit and
value-laden assumptions contemporary American culture appears to
make about avian attention. Finally, we describe a course in animal
cognition for undergraduates at the University of Utah which has
attempted to shed light on these contradictory positions and to
clarify their philosophical, psychological, and cultural origins. We
hope that this course sensitizes students to their own assumptions
about animal cognition and to the role of nonhuman animals in the
context of human culture. We optimistically hope also that learning
about multiple points of view, some of which may be sharply at odds
with their own strongly felt but perhaps not exhaustively examined
positions, will facilitate students’ making choices about their
own connections with nonhuman animals and that making these choices
will help them by their own personal standards to become more
responsible and constructive members of a society in which humans
and nonhuman animals interact in countless diverse ways.
Recent research on avian attention has developed mainly along the
lines previously laid out by the laboratory science of human
attention. We will therefore sketch this human research upon which
avian research rests. Before we do so, however, let us consider a
non-technical, in fact, unashamedly fanciful example of real world
nonhuman animal attention. We believe this kind of intuitive
example, described in plain English, can contribute to the
development of new experimental tasks and new theoretical
perspectives. Let us even permit ourselves, as non scientists often
do, to project our own thoughts onto the minds of nonhuman animals.
Suppose a seagull flies over a city park on a beautiful summer day,
above the heads of various groups of people eating a dizzying array
of food, ranging from hot dogs, hamburgers, and potato salad to
oysters Rockefeller, smoked salmon, and stuffed artichoke salad.
These foods appear to our seagull in a wide variety of visual
contexts, including in large and small uncovered and covered bowls
with shiny or dull, and transparent or opaque covers, and made of
porcelain, glass, plastic, or aluminum. Sometimes the bird sees food
in the hands of people and sometimes in small bits and crumbs
scattered about the lawn, on tables, or on blankets. Some food is
near to the seagull but in settings that might lead to defensive
actions by people if the bird were to try to get some for itself,
while other food is farther away but appears more accessible. Our
seagull is not the only nonhuman animal, or even the only bird on
the scene. There are dogs, ducks and geese; some walk near picnic
tables and groups of people, while others stand off at a distance.
Let us pretend that our seagull has a mental life in which it can
visually imagine defensive behaviors on the parts of these dogs and
birds as well as by the humans.
We would like to invite the reader to take every possible poetic
license and to imagine being the seagull, being hungry, and having a
long history of successfully seeing, obtaining, and eating human
food in situations like this. As you fly over the picnic grounds,
you see large patterns of food, including entire tables laden with
food and surrounded by large groups of people, and you see small
individual items of food, like a small piece of smoked salmon that
has fallen onto the lawn. You see all this food and you also see
lawn, parked cars, trees, large buildings in the distance, and
clouds in the sky. You also catch only quick glimpses of some foods,
as people open containers, put food onto their forks or into their
mouths, or as wind blows small crumbs about the park, while you get
prolonged sight of other foods lying on tables.
Is it possible that you, the seagull, might pay attention to
different kinds of environmental stimuli and events? Beyond our
fanciful, plain English story, is there any sense in which this is a
scientifically meaningful question? Our position is that all three
of the kinds of attention we will address below in detail may be
observed in this example. First, spatial attention might be in
evidence in that some regions in space, say those where food is most
plentiful, attracts more of the bird’s attention than other
regions of the park. Furthermore, we might suspect that as the bird
flies over the park, it redirects how different places attract more
or less attention. Perhaps the bird notices a quick flash of
sunlight on a package of popcorn in the hand of a young child as she
takes out a handful, and perhaps this happens so quickly as the bird
flies over that the bird has no chance even to move its head to see
the popcorn more clearly. Nevertheless, the bird notices the popcorn
in its container in a context that previously has on occasion
predicted the subsequent appearance of pieces of popcorn on the
ground. This brief stimulus might therefore be sufficient to cause
the bird to circle over the child. This is the question central to
spatial attention: How do different spatial locations associated
with important events or objects, or cues appearing in some spatial
locations but predicting important events in other spatial
locations, control how attention gets directed to those places?
Second, local/global attention is in evidence when the bird
sometimes pays greater attention to entire patterns of food, such as
an entire table spread with all sorts of different foods, and
sometimes pays greater attention to particular pieces of food, such
a particular bag of popcorn or a particular candy bar. The classic
defining exemplar of local/global attention is the familiar case
involving forests and trees. If a person is said "not to see
the forest for the trees," it is implied that the person is
paying attention, for some reason perhaps having to do with past
experience as well as current viewing conditions, to trees but that
under other conditions the person might pay attention to the forest.
In other words, it is implied that one can switch attention between
forest and trees. It is also implied that paying attention to either
forest or trees somehow facilitates processing of information about
the forest or trees, respectively. In our example, our seagull might
well from time to time switch between attending to groups of trees
and attending to individual trees.
Lastly, we might want to say that the bird selectively attends to
some features of complex stimuli more than to other features. For
example, the bird might pay greater attention to food than to cars
in the parking lot, or more to an unleashed bird dog than to a
chipmunk. We might intuitively assume that having paid more
attention to the dog than to the chipmunk the bird might then know
more about the dog than about the chipmunk: It might see the dog
more clearly and it might more accurately remember the dog. We might
wonder whether unattended stimuli would even be noticed or
remembered at all.
In general, researchers studying attention ask how knowing what
an organism pays attention to improves our understanding of the
organism’s behavior. The traditional assumption, which is not
without its problems but to which we will subscribe, is that there
is a limited amount of attention an animal has with which to respond
to its world; paying more attention to one thing means paying less
attention to other things. We assume that paying greater attention
to an event facilitates processing the corresponding information, so
that the bird will be faster, or more accurate, to respond to an
event to which it is paying greater attention, and it will
subsequently better remember something to which it has paid greater
attention. Because of the limited overall amount of attention a bird
is assumed to have, quicker responding to one stimulus might imply
slower responding to less well attended stimuli. These are classic
starting positions from which researchers study attention, knowing
full well that they are only assumptions and might be wrong not only
in individual cases but wrong in general as well.
Attention in Humans
Plain English found ample use for the idea of attention long
before there was a scientific psychology. When psychology emerged as
an academic discipline separate from philosophy and education,
little more than a hundred years ago, a pioneering attempt to come
to grips with the nature of attention was that of William James, who
assigned it a central role in his account of mental phenomena.
Within a generation, however, James’ view, and mentalistic
psychology in general, had fallen into disfavor due to the rise of a
behavioral approach that denied that mentalistic processes were
scientifically legitimate. More recently, Rorty (1980), Ryle (1949),
Skinner (1989), Wittgenstein (1953) and many others have clarified
conceptual and philosophical pitfalls in how natural language deals
with mental phenomena. For example, annoying conceptual confusions
can arise when one accidentally assumes through careless use of
visual metaphors based in natural language and popular culture, that
a homunculus, an imaginary little person inside the mind, causes a
person to attend to something. We respect a behavioral solution to
some of these problems and acknowledge the many pitfalls in the use
of mental terms. However, the behavioral approach is itself not
without its own set of problems, not the least of which is that our
scientific understanding of "behavior" may not be advanced
beyond that of "attention." Also, we find behaviorism’s
more restrictive language conventions sometimes heuristically
counterproductive as well as philosophically and scientifically
problematic (Shimp, 1976). Thus, in the present chapter, we permit
ourselves to use mentalistic terms, in part in the belief that they
facilitate communication with the nonspecialist to whom this chapter
is addressed, and in part in the belief that their heuristic power
motivates much or all of the laboratory research that in recent
years has revolutionized our understanding of avian attention. As
the present volume so clearly shows, we are not alone in these
beliefs (also see Allen and Bekoff, 1997; Ristau, 1991). Perhaps
most importantly, we suspect that the language of
"attention" will facilitate a dialog between scientific
and popular accounts of the relation between human and nonhuman
animal attention, which is our chief purpose in this chapter.
Let us now look at three examples of the kinds of naturalistic
behaviors in humans which in natural language invoke the idea of
attention. These examples are spatial attention, local/global
attention, and selective attention. Let us look at each in turn.
Spatial attention. Intuition leads us to believe that people
do not simultaneously attend equally to all locations in their
environments. Some locations are more important than others, and
people may attend more to those locations than to others. Imagine,
for example, a situation in which the driver of a car rapidly
approaches a dangerous intersection where there is cross traffic
from both left and right, and especially heavy traffic often
appearing from the left. The driver might be aware that some
drivers, even though cross traffic has a stop sign, do not stop and
that fatal accidents have resulted. The driver might therefore keep
her eyes focused on the road ahead and yet also pay more attention
than she otherwise would to peripheral spatial locations on the left
from which traffic is likely to appear.
Posner and his colleagues (Posner, 1980) and many others have
developed experimental methods and concepts by which this kind of
attention, spatial attention, can be studied. A common metaphor has
been that spatial attention is like a mental spotlight, which swings
toward an important location and "illuminates" that
location in the mind’s image of what either has already happened
there or is expected shortly to happen there. It is assumed that
when the spotlight illuminates a location in a mental image, an
event occurring in the actual corresponding location is processed
more quickly or more accurately, so that the spotlight improves
processing of events in that location. Posner’s method instructs a
person to respond quickly and accurately to target stimuli that
appear in either of two locations. Just before a target appears, a
priming cue appears. This cue may probabilistically predict the
location of the subsequent target. When it correctly predicts the
target location on a trial, the trial is called a valid trial. An
invalid trial occurs when predicted target location and actual
target location are different. Typically, targets are arranged in
the predicted location with a probability, called cue validity, that
is much greater than the probability with which the target appears
in the other, non cued, location. The "validity effect"
occurs when reaction times to respond to targets are faster on valid
trials than on invalid trials; humans typically show this validity
effect, which is to say, they use the probabilistic information in
priming cues to anticipate the location of the subsequent target.
Metaphorically speaking, they swing their attentional spotlights
toward spatial locations they judge likely to be important. The
careful driver of the car approaching the dangerous intersection
would be said to move her attentional spotlight to the left, while
keeping her eyes on the road ahead.
Let us consider some additional details of human spatial
attention in order both to understand better how it is studied with
human subjects and to establish a basis for comparison when we later
describe roughly analogous nonhuman animal spatial attention.
Consider a few aspects of an experiment by Posner, Nissen, &
Ogden (1978). They showed human subjects a target letter X in either
of two spatial locations, to the left or right of a central box at
which subjects fixated. A subject was asked to press a left key as
quickly as possible when the target appeared on the left, and a
right key when the target was on the right. A warning signal
appeared shortly before the target appeared. This warning cue was
either a plus sign, an arrow pointing to the left, or an arrow
pointing to the right. The plus sign appeared on a random half of
the trials and was neutral in the sense that it had a cue validity
of .50, so that it did not predict the location of the subsequent
target better than chance. The arrow pointing to the left, however,
had a cue validity of .80, so that 80 % of the time when it
appeared, it pointed in the direction of the subsequent target, and
correspondingly for the arrow pointing to the right. The length of
time between the warning cue and the target was varied and was
either 0 (no warning cue), or 50, 150, 300, 500, or 1000 msec. All
told, several hundred trials were presented. Figure 2 (which is
taken from Figure 1 in Posner et al., 1978, page 141) shows average
reaction time to press the correct key as a function of the time between
the warning 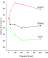 cue and the target. (See Figure 2) The top, middle, and bottom curves
are for conditions for which the cue validity was .2, .5., and .8,
respectively. For present purposes, the two chief results are that
1) reaction times were fastest for valid cues (bottom curve), less
fast for neutral cues (middle curve), and slowest for invalid cues
(top curve), and 2) this validity effect persisted for at least one
sec. Thus, humans direct attention to spatial locations where
important events are expected to occur, and this effect persists for
at least some short period of time. Our imaginary example of a
driver approaching a dangerous intersection, which makes perfectly
good intuitive sense, therefore also makes good sense from the
perspective of laboratory research on spatial attention.
cue and the target. (See Figure 2) The top, middle, and bottom curves
are for conditions for which the cue validity was .2, .5., and .8,
respectively. For present purposes, the two chief results are that
1) reaction times were fastest for valid cues (bottom curve), less
fast for neutral cues (middle curve), and slowest for invalid cues
(top curve), and 2) this validity effect persisted for at least one
sec. Thus, humans direct attention to spatial locations where
important events are expected to occur, and this effect persists for
at least some short period of time. Our imaginary example of a
driver approaching a dangerous intersection, which makes perfectly
good intuitive sense, therefore also makes good sense from the
perspective of laboratory research on spatial attention.
Local/Global attention. Spatial locations are not the only
environmental features that attract attention. Plain English
suggests that one can attend either to an entire object or to a part
of an object. Gestalt psychology made much of this same idea, in the
form of perception of wholes or parts, as in, "He cannot see
the forest for the trees." Navon, Robertson,  Lamb, their
colleagues, and others, have developed experimental methods and
theory by which this kind of attention, called local/global attention, can be studied. (See Figure 3) In such an experiment, a human
participant is usually instructed to respond as quickly as possible
to a target stimulus that appears at either the local or global
level of a compound alphanumeric stimulus, such as in Figure 3. Lamb, their
colleagues, and others, have developed experimental methods and
theory by which this kind of attention, called local/global attention, can be studied. (See Figure 3) In such an experiment, a human
participant is usually instructed to respond as quickly as possible
to a target stimulus that appears at either the local or global
level of a compound alphanumeric stimulus, such as in Figure 3.
In the case represented in the bottom half of Figure 3, a person
might be instructed to respond to a left button when she sees the
letter H and to respond to the right button when she sees the letter
S. A global letter is composed of many small local letters. The
letter H might appear either as a large, global letter composed of
many, smaller instances of some other letter, such as many small X’s,
or as a small, local letter many of which compose some other global
letter, such as T. Similarly, the letter S might appear either as a
global letter composed of other local letters or as the smaller
local letter many of which form a larger global letter. In a typical
experiment, a target letter occurs on any one trial at only one
level and an irrelevant distractor stimulus occurs at the other
level. Over trials, a target letter appears at both levels. It has
been found that people can be primed to expect targets at a
particular level, in the sense that when most targets appear at a
particular level, so that targets at that level become more expected
than targets at the other level, or when a priming stimulus precedes
a target and probabilistically predicts a target at a particular
level, targets at that primed level are processed more quickly
and/or accurately (Lamb & Robertson, 1990; Robertson, 1996).
Thus, a basketball fan might be said to be primed to attend to a
local level when he clearly sees Karl Malone’s steal of the ball
from an opponent, immediately after the fan’s wife draws his
attention to Malone, who is a local feature in comparison to the
entire pattern of players on the floor. Similarly, the fan might be
primed to attend to a global level after his wife mentions that a
team has just gone to a different defensive pattern, a global
feature characteristic of the entire pattern of players, after which
he fails to notice a similar steal by Malone, a local feature.
Selective attention. The "cocktail party effect" is
a common illustration of selective attention; a guest at a cocktail
party who focuses attention on the conversation in which she is
participating while simultaneously ignoring, and consequently
feeling unaware of other conversations in the same room, is still
able to switch the focus of her attention from the current
conversation to another, particularly if some important piece of
information, such as her own name, is mentioned in an unattended
conversation. Many early models of selective attention focused on
this effect and characterized attention as a selective filter,
allowing some environmental information to be processed further
while filtering out other, less useful information. Such a filtering
mechanism may be invoked when it is assumed that the mind has
limited processing resources. This limitation has been seen as
implying a need to select important information for processing,
while ignoring relatively unimportant information and thereby
conserving cognitive resources. One common method of demonstrating
selective attention in the laboratory is the dichotic listening
task, where different information is presented simultaneously to a
participant’s left and right ears. Participants have often been
shown to attend to information in one channel much better than to
that in the other. Thus, selective attention is demonstrated when
participants can attend to one source of information, while
ignoring, and remaining relatively unaffected by, the other. The
cocktail party effect is almost a defining exemplar of this process
of selective attention.
Selective attention has also been demonstrated in many other ways
in humans, including in almost countless experiments using visual
instead of auditory stimuli. We choose here to demonstrate the
phenomenon in terms of a "randomization" categorization
task developed by Ashby, Gott, Lee, Maddox, and others, to study
"ill-defined" categories in humans (see Chase
and Heinemann 2001; Huber, 2001;
Urcuioli, 2001).
"Ill-defined" categories would intuitively seem a poor
choice if one hoped to obtain a clear, unambiguous understanding of
the categorization process. Indeed, such categories have the
seemingly unpleasant characteristic of not having any single feature
or set of features by which one can unambiguously determine the
category to which an exemplar belongs. Why would anyone study
ill-defined categories, when they could study tidy, logical, and
generally agreeable well-defined categories? The answer is that
real-world categories may seldom, if ever, be well defined (Douglas
and Hull, 1992; Rosch & Mervis, 1975; Wittgenstein, 1953), and
ill-defined categories permit the evaluation of a wider range of
theories of categorization.
In a typical version of the randomization task, a participant
categorizes a series of two-dimensional stimuli as belonging to one
or the other of two ill-defined categories. The categories are ill
defined in the sense that there is no feature or combination of
features that guarantees membership in a category. Indeed, in the
task we describe, the very same stimulus can at times belong to one
category and at other times to the other, rather like a Gestalt
reversible figure, that can be seen now as a vase and then as two
human faces directed toward one another, as in Figure 1 above
(Wittgenstein, 1953).
Figure 4 (which is taken from Figure 1 in Herbranson, Fremouw,
and Shimp, 1999, page 114) provides a way of looking at the nature
of the randomization task. This Figure is actually from an
experiment with pigeons described below, yet serves to summarize the
corresponding human procedures which are our focus in this Section. (See Figure 4)
The top panel shows two ill-defined categories, A and B, in the form
of two overlapping normal bivariate distributions. The distributions
are drawn in what is referred to as the "stimulus space,"
in which each point corresponds to a specific two-dimensional
stimulus, such as a rectangle. The horizontal and vertical
coordinates of a point correspond to the width and height of a
rectangular stimulus and the third coordinate corresponds to the
probability with which the stimulus is shown to a subject given a
particular category. The top panel shows two equal-likelihood
contours corresponding to points that are equally likely to occur
given a particular category. The bottom panel shows two such
equal-likelihood contours along with a diagonal line which is the
optimal decision bound, the set of points equally likely to occur
given either category. Optimal performance by a subject in the
divided attention, or "tall/wide" task consists of
categorizing rectangles corresponding to points above the line as
belonging to category A and of categorizing rectangles corresponding
to points below the line as belonging to category B.
(See Figure 4)
The top panel shows two ill-defined categories, A and B, in the form
of two overlapping normal bivariate distributions. The distributions
are drawn in what is referred to as the "stimulus space,"
in which each point corresponds to a specific two-dimensional
stimulus, such as a rectangle. The horizontal and vertical
coordinates of a point correspond to the width and height of a
rectangular stimulus and the third coordinate corresponds to the
probability with which the stimulus is shown to a subject given a
particular category. The top panel shows two equal-likelihood
contours corresponding to points that are equally likely to occur
given a particular category. The bottom panel shows two such
equal-likelihood contours along with a diagonal line which is the
optimal decision bound, the set of points equally likely to occur
given either category. Optimal performance by a subject in the
divided attention, or "tall/wide" task consists of
categorizing rectangles corresponding to points above the line as
belonging to category A and of categorizing rectangles corresponding
to points below the line as belonging to category B.
A common way to implement the randomization task is to define the
two dimensions along which stimuli vary as the widths and heights of
rectangles or of other geometric figures a human participant
observes. Two important versions of the task are the
"tall/wide" or divided attention task and the "one
dimension" or selective attention task. Let us consider each in
turn. In the tall/wide task, a rectangle taller than it is wide is
optimally categorized as belonging to one category, and a rectangle
wider than it is tall is optimally categorized as belonging to the
other. (An optimal categorization is a choice of the response
alternative with the higher probability of reinforcement.) If a
participant optimally categorizes individual rectangles, then the
participant may be said to divide attention between both dimensions,
because optimal categorization requires integrating information from
both dimensions.
Figure 5 (which is taken from Figure 2 in Herbranson et al.,
1999, page 117) shows greater detail for the task involving divided
attention. The Figure shows the degree to which the two ill-defined
categories overlapped: The prototypes or average rectangles for each
category are shown as heavy points in the center of two
equally-likely contours, one drawn at one standard deviation from
the prototype and one at two standard deviations. The bottom panel
of Figure 5 shows the actual prototypes for the experiment.
Consider an experiment that showed humans perform in nearly
optimal ways on this task (Ashby and Gott, 1988). In their
Experiment 1, three humans viewed stimuli on a computer monitor. The
stimuli consisted of two lines, one vertical and the other
horizontal, joined at the upper left corner; the stimulus contained
a corner and two  edges of a rectangle. (See Figure 5)
Each stimulus belonged either
to category A or to category B. Each category had a corresponding
bivariate distribution that was approximately normal. The subjects
saw exemplars of each category equally often, on the average, and on
any particular trial, the category was chosen by essentially tossing
an unbiased coin, and then the stimulus was obtained by randomly
selecting a sample from the corresponding bivariate distribution. A
subject viewed each stimulus and categorized it as belonging to
either category A or B by pressing buttons that corresponded to the
categories, as in left for A and right for B. At the end of each
trial, a participant was informed to which category the stimulus on
that trial actually belonged. Each of the participants was given a
few hundred training trials. For present purposes, we wish to
examine only one of the several important theoretical questions
Ashby and Gott analyzed.
edges of a rectangle. (See Figure 5)
Each stimulus belonged either
to category A or to category B. Each category had a corresponding
bivariate distribution that was approximately normal. The subjects
saw exemplars of each category equally often, on the average, and on
any particular trial, the category was chosen by essentially tossing
an unbiased coin, and then the stimulus was obtained by randomly
selecting a sample from the corresponding bivariate distribution. A
subject viewed each stimulus and categorized it as belonging to
either category A or B by pressing buttons that corresponded to the
categories, as in left for A and right for B. At the end of each
trial, a participant was informed to which category the stimulus on
that trial actually belonged. Each of the participants was given a
few hundred training trials. For present purposes, we wish to
examine only one of the several important theoretical questions
Ashby and Gott analyzed.
Specifically, let us ask if humans were able to learn to
categorize two-dimensional stimuli in nearly optimal ways. Figure 6
suggests an answer. Figure 6 shows how one person  categorized the
stimuli on 300 individual trials. (See Figure 6) Categorization of a stimulus as
belonging to category A or B appears as an "x" or as a
filled circle, respectively. Figure 6 shows that the great majority
of stimuli categorized as belonging to A are above the straight line
with a slope of one and the great majority of stimuli categorized as
belonging to B are below this line. Thus, stimuli taller than wide
were usually categorized as A and stimuli wider than tall were
usually categorized as B. This is the outcome roughly corresponding
to the optimal solution represented by the straight line with slope
of one. (See Ashby and Gott (1988) for details.) The person
for whom results are displayed in Figure 6 therefore appeared to
have used information from both dimensions of stimuli, height and
width, and integrated this information in a nearly optimal way. This
person therefore may be said to have divided attention between the
two dimensions of height and width.
categorized the
stimuli on 300 individual trials. (See Figure 6) Categorization of a stimulus as
belonging to category A or B appears as an "x" or as a
filled circle, respectively. Figure 6 shows that the great majority
of stimuli categorized as belonging to A are above the straight line
with a slope of one and the great majority of stimuli categorized as
belonging to B are below this line. Thus, stimuli taller than wide
were usually categorized as A and stimuli wider than tall were
usually categorized as B. This is the outcome roughly corresponding
to the optimal solution represented by the straight line with slope
of one. (See Ashby and Gott (1988) for details.) The person
for whom results are displayed in Figure 6 therefore appeared to
have used information from both dimensions of stimuli, height and
width, and integrated this information in a nearly optimal way. This
person therefore may be said to have divided attention between the
two dimensions of height and width.
Selective attention may also be demonstrated with this
randomization procedure. This version of the task uses the same set
of stimuli but uses different sampling distributions to define the
two ill-defined categories. In this task, rectangles continue to
vary from trial to trial in terms of both width and height but only
a single dimension,  say width, is relevant to the definition
of the
categories. (See Figure 7) Let category A consist of angles the widths of which are
greater than some specific width, and let the other category consist
of angles the widths of which are less than this same value. Let
height vary from stimulus to stimulus as before, but let it be
irrelevant to whether a stimulus belongs to category A or B. To
perform optimally in this task, a participant needs to categorize
exclusively on the basis of width and to ignore height. Figure 7
shows the stimulus space and optimal decision bound for such a task. say width, is relevant to the definition
of the
categories. (See Figure 7) Let category A consist of angles the widths of which are
greater than some specific width, and let the other category consist
of angles the widths of which are less than this same value. Let
height vary from stimulus to stimulus as before, but let it be
irrelevant to whether a stimulus belongs to category A or B. To
perform optimally in this task, a participant needs to categorize
exclusively on the basis of width and to ignore height. Figure 7
shows the stimulus space and optimal decision bound for such a task.
It is of course possible to switch dimensions in such a task and
to make height relevant and width irrelevant. Figure 8 shows the
results from one human performing on such a task, with the criterion
height indicated by the horizontal line. Figure 8 shows that the
person did in fact strongly tend to categorize angles shorter than
the criterion height as belonging to one category and taller than
the criterion as belonging to the other category. (See Figure 8) Thus, the person
selectively attended to the relevant height dimension and ignored
the irrelevant width dimension. The randomization task has been used
to show that humans often closely approximate the optimal solution
in this and other versions of the randomization task (Ashby and Gott,
1988; Ashby and Maddox, 1998).
category. (See Figure 8) Thus, the person
selectively attended to the relevant height dimension and ignored
the irrelevant width dimension. The randomization task has been used
to show that humans often closely approximate the optimal solution
in this and other versions of the randomization task (Ashby and Gott,
1988; Ashby and Maddox, 1998).
These three examples of spatial attention, local/global
attention, and of divided and selective attention, illustrate just a
few of the ways in which human attention can be scientifically
studied and how laboratory research can clarify our intuitive ideas
about attention. These examples are selected to enable a reader to
appreciate better the parallels we now wish to draw between human
attention and nonhuman animal attention.
Avian Attention in the Laboratory
The idea that a nonhuman animal can "attend" to
anything at all has a checkered past in classical philosophy and
Western thought. Contrast the sympathetic position of Saint Francis,
who so often is portrayed with ostensibly happy birds flying about
his head and perched on his shoulders, with that of Descartes, who
is reputed to have laughed at the sounds emitted by animals upon
which he was operating without anesthetics. His laughter derived
from his thorough belief that animals were machines and from the
resulting laughable absurdity in the similarity between human cries
of pain and the sounds emitted by his animal machines. This
Cartesian dualism continues to play an important, but not
correspondingly explicit, role in contemporary culture and science,
in part perhaps because Descartes’ position was inspired by
theological issues that are rarely discussed in scientific circles.
The animal mind initially was seen as a legitimate and important
part of the science of animal behavior (Romanes, 1898). The idea
from Darwinian evolutionary theory, that there should be some form
of mental continuity across species very roughly corresponding to
anatomical continuity, was adequate motivation for early animal
behaviorists to search for indications of animal intelligence,
including attention. They found what they were looking for.
This initial outburst of enthusiasm for the scientific study of
mental life in animals was quickly suppressed, however, by
consequences of inadequate scientific skepticism: Early in the 20th
century, for example, "Clever Hans," a horse touted to be
able to count and communicate with its trainer by appropriately
tapping its hoof, was found instead to rely only on unintentional
gestures of humans who knew when the horse should stop tapping (Candland,
1993). A conservative reaction set in that suppressed not only the
use of anecdotal evidence based on natural language and culture, but
the development of empirical methodology by which complex behaviors
could be scientifically investigated For subsequent generations, it
was an uphill scientific battle to investigate a cognitive process
in a nonhuman animal. The usual presumption was, and still is, that
the virtue of parsimony is on the side of "simple"
explanations in terms of a non cognitive, purely mechanistic,
behavioral position (Shimp, 1999). For example, when the concept of
memory was explicitly advocated as a legitimate concept in the
experimental analysis of animal behavior (Shimp, 1976), it caused
explicit discontent (Branch, 1977), and researchers studying
short-term memory often had to show that a "simple" linear
chaining, associative learning account failed before they were
permitted to interpret results in terms of memory. Correspondingly,
when one now wishes to interpret phenomena in terms of the concept
of attention, one often must first show that an account in terms of
memory is not plausible. Thus, what is simple has changed; first, it
was conditioning, then it was expanded to include memory, and now,
attention seems to be becoming part of an acceptably parsimonious
system. "Simplicity" itself is anything but simple
(Hacking, 1999; Goodman, 1977), the criteria for determining what
"simple" means are virtually never made explicit (Shimp,
1999), and even when they are, their appropriate use may be
contingent on many contextual assumptions that remain only implicit
(Hanson, 1969; Goodman, 1977).
Even during the period when behaviorism defined the conventional
evaluative standards, there were some forcefully expressed opposing
views (Krechevsky, 1932; Tolman, 1932), and a rather solitary paper
by Reynolds (1961) suggested that the notion of attention need not
threaten a behavioral perspective. With the establishment of a
science of animal cognition in the 1970's, however, it became much
more acceptable in some circles to discuss attention or
attention-like phenomena in nonhuman animals (Kamin, 1969; Riley and
Roitblat, 1978). The chapter by Sutton &
Roberts (2001) in the present
volume provides a brief summary of this stage of conceptual and
methodological development.
At about the same time, the idea of a "search image"
became established within ethology as an explanation for why
foraging animals acquire temporary biases in favor of prey that
locally appear more frequently (Tinbergen, 1960): This ethological
idea is similar to the cognitive idea that a predator adapts to
local contexts by learning to attend to, or look for, a type of prey
which temporarily is more likely to be found (Fremouw, et al.,
1998).
The present volume attests to the many forms of avian attention
that now can be studied as legitimate scientific concepts. We focus
on three of these, corresponding to the three types of human
attention reviewed above.
Spatial attention. Our research on spatial attention was conducted in collaboration
with Dr. Frances Friedrich, whose own research involves, among other
things, neuropsychological applications of the Posner paradigm for
the study of spatial attention in humans. To see if some of the
basic phenomena of spatial attention could be obtained in nonhuman
animals, we selected pigeons as the experimental participants
because they had already been shown to possess many human-like
visual capabilities and they seemed likely candidates for having
developed ways to deal with how important environmental events occur
rapidly in predictably different spatial locations. We set out to
see if we could obtain the validity effect described previously.
Our method (Shimp and Friedrich, 1993) generally paralleled that
described above for humans but with a few important differences. A
pigeon’s task was to peck an illuminated left or right target key
the location of which varied randomly from trial to trial. A peck to
the target key delivered food. Immediately before the left or right
target key was illuminated, a very brief (50 msec) white light
flashed on either the left or right key. Like Posner et al., (1978),
we were interested in the persistence of any spatial attention we
might discover, so like them, we varied the time between the
occurrence of the priming cue and when the target appeared. This
priming cue had a cue validity of .80. That is, 80% of the time,
when the cue flashed on the left, the target subsequently appeared
on the left, and 80% of the time when the cue flashed on the right,
the target subsequently appeared on the right. The location of the
cue, in other words, was a reasonably reliable indicator of where an
important environmental event, the target, was about to occur.
Notice that in the case of the experiment by Posner et al., (1978),
the priming cue was a central arrow pointing either to the left or
right. In our experiment, the priming cue occurred in the actual
spatial location at which targets appeared. This difference between
central symbolic cues and cues occurring in the left or right
spatial locations is important in the human literature, but symbolic
cues have yet to be studied extensively in the case of avian spatial
attention. Our basic question was whether valid cues, those
appearing in the same locations as the subsequent targets,
facilitated processing targets in those locations, in comparison to
invalid cues, which appeared in the locations opposite to those of
targets.
The answer was, Yes. Figure 9 shows reaction time as a
function of the time between cue and target. The figure shows that
reaction times to respond to targets preceded by valid cues (bottom
curve) were faster than reaction times to respond to targets after
invalid cues (top curve), and this validity effect persisted for
some duration of time. 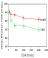 (See
Figure 9) This is the chief defining feature of spatial
attention, so that we now know that pigeons, as well as humans,
display that attentional phenomenon. While not all the outcomes were
exactly like those from humans, the parallels were amply close to
justify continued examination of mental continuity between nonhuman
animal and human spatial attention. Subsequently, spatial attention
has been demonstrated also in rats (Bushnell, 1995). (See
Figure 9) This is the chief defining feature of spatial
attention, so that we now know that pigeons, as well as humans,
display that attentional phenomenon. While not all the outcomes were
exactly like those from humans, the parallels were amply close to
justify continued examination of mental continuity between nonhuman
animal and human spatial attention. Subsequently, spatial attention
has been demonstrated also in rats (Bushnell, 1995).
Local/Global
attention.
We again chose pigeons to explore whether a nonhuman animal can
display a human perceptual phenomenon, in this case, shifts of
attention between local and global levels of perceptual analysis.
Figure 3 displayed above in Section II shows the stimuli we used.
We arranged compound stimuli on computer monitors in experimental
chambers in which the pigeons performed (Fremouw, et al., 1998).
Each stimulus a pigeon viewed had two "levels," a local
and a global, and on each trial, a target appeared at one or the
other level. If a bird had targets consisting of the letters H and
S, these could appear either as global letters made up of local
distractors (irrelevant characters never appearing as targets) or as
local letters forming a global distractor. If H appeared on a trial,
a bird got food if it pecked the left key, and if S appeared, it got
food if it  pecked the right key. Distractor letters were T and E.
Both targets, H and S, appeared equally
often (click here see an interactive
demonstration of this procedure). The level at which
they appeared, however, was systematically varied over conditions,
in some of which targets appeared on a randomly selected 85% of the
trials at the global level and targets at the local level appeared
the other 15% of the time. Every few days, a condition was changed
so that the level at which targets appeared more frequently was
reversed. We asked if, as in the corresponding human experiments,
biasing targets at a particular level facili pecked the right key. Distractor letters were T and E.
Both targets, H and S, appeared equally
often (click here see an interactive
demonstration of this procedure). The level at which
they appeared, however, was systematically varied over conditions,
in some of which targets appeared on a randomly selected 85% of the
trials at the global level and targets at the local level appeared
the other 15% of the time. Every few days, a condition was changed
so that the level at which targets appeared more frequently was
reversed. We asked if, as in the corresponding human experiments,
biasing targets at a particular level facili tated an animal’s
processing of targets at that level, as reflected in faster
responding to those targets. Figure 10 shows that birds did in fact respond faster to local
targets than to global targets when targets appeared more frequently
at the local level. Similarly, reaction time was faster to global
targets than to local targets when targets appeared more frequently
at the global level. We found that the familiar process of human
local/global attention obtains also in nonhuman animals (however,
see Cook's chapter (2001) for a different set of results
with similar
stimuli). Thus,
local/global attention, like spatial attention, is not a purely
human phenomenon, and the concept of mental continuity is once again
strengthened. (see Figure 5) tated an animal’s
processing of targets at that level, as reflected in faster
responding to those targets. Figure 10 shows that birds did in fact respond faster to local
targets than to global targets when targets appeared more frequently
at the local level. Similarly, reaction time was faster to global
targets than to local targets when targets appeared more frequently
at the global level. We found that the familiar process of human
local/global attention obtains also in nonhuman animals (however,
see Cook's chapter (2001) for a different set of results
with similar
stimuli). Thus,
local/global attention, like spatial attention, is not a purely
human phenomenon, and the concept of mental continuity is once again
strengthened. (see Figure 5)
Selective attention.
There are many ways to demonstrate selective attention in
nonhuman animals. In this case, we again chose pigeons for their
visual capabilities. We demonstrated the phenomenon of selective
attention in terms of the randomization categorization task
described above for humans. It will be recalled that two common
versions of this task are the integration version, or the
"tall/wide" version that requires an individual to
integrate information from two characteristics of a complex
stimulus, which is the divided attention version, and the "one
dimension" or selective attention version. In our experiments,
we showed pigeons rectangles, and in the tall/wide task, a pigeon
optimally categorized a rectangle taller than it was wide by pecking
the left key, representing one category, and optimally categorized a
rectangle wider than it was tall by pecking the right key,
representing the other category.
Figure
5 represents our specific version of
the "tall/wide" task (click here see an interactive
demonstration of this procedure) . Figure 11 shows the results for
one of four birds. This particular bird was the "best" in
a technical sense beyond the scope of the present brief summary.
(See
Figure
11) Recall that optimal categorization would lead to a bird’s calling
rectangles represented by points above the bold diagonal line with a
slope of one exemplars of one category, and rectangles represented
by points below the line as sense beyond the scope of the present brief summary.
(See
Figure
11) Recall that optimal categorization would lead to a bird’s calling
rectangles represented by points above the bold diagonal line with a
slope of one exemplars of one category, and rectangles represented
by points below the line as  exemplars of the other category.
Individual points and circles represent a bird’s individual
categorization responses. Figure 11 shows that the bird’s
responses tended in general to conform to the optimal rule. We
discovered that pigeons, like humans, divide attention between the
two dimensions and perform in nearly optimal ways on this task (Herbranson,
et al., 1999). exemplars of the other category.
Individual points and circles represent a bird’s individual
categorization responses. Figure 11 shows that the bird’s
responses tended in general to conform to the optimal rule. We
discovered that pigeons, like humans, divide attention between the
two dimensions and perform in nearly optimal ways on this task (Herbranson,
et al., 1999).
The selective attention version of our task used the same set of
stimuli but used different sampling distributions to define the two
ill-defined categories. Figure 7
displayed above in Section II
describes the specific arrangements for our selective attention
task. In this task, rectangles continue to vary from trial to trial
in terms of both width and height but only a single dimension, say
width, is relevant to the definition of the categories. One category
might consist of rectangles the widths of which are smaller than a
particular value, and the other category would consist of rectangles
the widths of which are greater than this value. Thus, to perform
optimally in this task, a pigeon needs to categorize exclusively on
the basis of width, and to ignore height, which it will be recalled,
continues to vary from trial to trial. Figure 12 shows the
results for one of four birds in this version of the
randomization
task. (See Figure 12) Again, the Figure shows the results from the bird that
satisfied a technical criterion to the best degree. The Figure shows
that the bird categorized rectangles in rough agreement with the
optimal rule represented by the bold vertical line, and accordingly,
it may be said to have selectively attended to the dimension of
width and to have ignored the dimension of height. One bird, not
shown, did not conform to this rule. In general, however, taking all
four birds into account, as in the case for divided attention, we
found that pigeons, like humans, approximate the optimal solution in
the selective attention version of the randomization task (Herbranson
et al., 1999). Figure 12 shows the
results for one of four birds in this version of the
randomization
task. (See Figure 12) Again, the Figure shows the results from the bird that
satisfied a technical criterion to the best degree. The Figure shows
that the bird categorized rectangles in rough agreement with the
optimal rule represented by the bold vertical line, and accordingly,
it may be said to have selectively attended to the dimension of
width and to have ignored the dimension of height. One bird, not
shown, did not conform to this rule. In general, however, taking all
four birds into account, as in the case for divided attention, we
found that pigeons, like humans, approximate the optimal solution in
the selective attention version of the randomization task (Herbranson
et al., 1999).
Implications for "Mental
Continuity"
We have displayed three similarities between human and avian
attention. These similarities are what late 19th century
and early 20th century animal behaviorists and other
advocates of Darwinian mental continuity were looking for. Early
animal behaviorists lacked the necessary scientific methods and
concepts by which to demonstrate mental continuity to the
satisfaction of skeptics who saw nonhuman animals in purely
mechanical terms. The modern experimental methodology we have
described seems to provide the necessary tools and seems to satisfy
many, if not all (Budiansky, 1998; Kagan, 1998) observers, about the
possibility of nonhuman animal cognition. Of course, those
researchers who feel that any language that includes terms like
"attention" is misguided would wish to rephrase all of the
preceding discussion of attentional processes. The discussion could
be rephrased in terms of shaping, stimulus control, discrimination
learning, generalization, reinforcement contingencies, and so on.
For the reasons we described previously, while we respect the
reasons why some individuals would want to use a different language
for research on attention, we believe that on the whole a
"cognitive" or even mentalistic language is more helpful.
We believe these three examples of similarities between human and
avian attention imply a duty to look for still more evidence for
mental continuity corresponding to continuity of biological
structure and function. Just as the natural world encourages some
continuity in anatomical and neuroanatomical structure, and in
processes of digestion, locomotion, and so on, it appears to
encourage certain forms of continuity in cognitive functioning.
The Sky is the Limit
The very success of this volume in portraying an abundance of
similarities between human and avian visual cognition might mislead
a reader into thinking much more is known about avian cognition than
seems to us to be the case. The sky is still the limit for what
there is yet to be learned about the avian mind. We doubt that the
science has even begun to identify some of what the future will show
are many of the most exciting questions. Little is known about the
nature of avian mental life in comparison to results from human
neuropsychology, research on aware versus unaware processing, or
metamemory. A very small beginning has been made on awareness, self
awareness, metaknowledge, and other matters related to the nature of
memory (Clayton and Dickinson, 1998, 1999; Inman and Shettleworth,
1999; Shimp, 1982, 1983; Shimp, Fremouw, Ingebritsen, and Long,
1994), but the difference in progress between research on these
issues in humans and nonhumans is pronounced. We think this lack of
knowledge teaches humility. It also sets the stage for the rest of
this chapter. If we knew just what a pigeon was seeing, thinking,
and feeling, we would feel better informed as to what appropriate
cultural connections there could be or should be between humans and
pigeons. Absent much information about who, so to speak, a pigeon
is, it is exceptionally difficult for us to know how we should
interact with it.
The Animal Mind and the University
Curriculum: Historical Trends
If not even behavioral scientists researching the avian mind know
"who a pigeon is," it is scarcely surprising that college
students are conflicted about the avian mind. Students sometimes see
birds, and animals in general, especially their own pets, as highly
similar to humans, and in this sense, they take a very strong
position on the idea of mental continuity; students at these times
are prone to see animals as having mental lives much like their own,
and may believe they share an empathic understanding with their
pets. Some students at other times seem less interested in, and
concerned about, nonhuman animal mental lives, and seem instead to
adopt a more Cartesian view that nonhuman animals are essentially
mechanical and are irrelevant to questions about human mental lives.
We have already seen above how these same differences in student
opinions about the animal mind have been reflected in different
scientific views about the animal mind. How the animal mind has been
presented over the past few decades in classes in animal learning,
conditioning, and cognition at the University of Utah has reflected
many of these differences. Emerging trends can be seen in how these
classes have evolved, and if we speculate and extrapolate these
trends, we may glimpse important features of the future of avian
visual perception and of animal cognition.
Consider an example which, though hypothetical and
oversimplified, captures some of the pedagogical challenges which
over the years the course on learning and conditioning has faced.
Many students have pets. Let us imagine that a student has a pet
parrot, Hector. She loves Hector, and often wonders what Hector is
thinking and feeling. She is a psychology major, and when she spies
a course on animal learning listed in the university catalog, she
signs up, hoping vaguely that, among other things, she might learn
something about her pet bird. Let us speculate how such a student
might have reacted to various versions of the course at the
University of Utah that have been offered by several different
instructors in the past 35 years or so.
Psychology classes in conditioning and learning in the 1960's
generally presented an impersonal, value-free and positivistic
picture of science. In this context, Cartesian dualism was easy for
a student to understand. In a word, a bird was a machine, despite
all intuitive appearances to the contrary. From this perspective,
anything apparently mental was unscientific. A student was not
encouraged to think about the relevance of the course to her pet
except in terms of conditioning or the shaping of behavior, such as
control of a parrot’s behavior through reinforcement of squawking
at or intimidating people. The problem was that her pet belonged to
the real world, which by the reductionistic standards of laboratory
science, was messy and excessively complex. According to this view,
the laboratory was the place to understand legitimate approaches to
the animal "mind." and a student quickly learned to ignore
plain English and the concept of the mind as not only entirely
unscientific but as inimical to the development of science. In
particular, the notion of "attention" was viewed with such
suspicion by the laboratory science of nonhuman animal behavior that
the term usually did not appear in undergraduate texts or the
scientific literature. The student left the class having learned
technical material about learning and conditioning, with some
practical information deriving from behavior analysis. The student
learned little about the emotional or perceptual life of her pet,
and may have acquired the habit of not considering empathic
understanding as relevant to a scientific understanding of her pet.
She learned a rather forbiddingly technical behavioral language, and
learned also that to talk in plain English about her pet’s mental
life was forbidden. The student might have left the class thinking
the science of animal learning had little or nothing to say about
the very behaviors of her pet that meant the most to her. In this
sense, science and everyday life seemed deeply at odds with each
other: They not only dealt with different features of an animal’s
life, they did so in a way that often seemed to involve
disparagement by scientists of students’ intuitive ideas about
animal minds.
The 1970's saw here and there a mild loosening of the ban on
cognitive terminology, especially regarding the acceptability of
"memory." A growing realization of the relevance of
evolutionary biology, and therefore of real world animal behavior,
contributed to the understanding of basic learning processes and
contributed also to a growing tolerance for more complex behavior,
such as foraging and caching. Nevertheless, despite an emerging
trend to explore mental continuity across species, the science
tended on the whole to focus on elementary laboratory behaviors, and
our imaginary student might not have felt much differently about the
relevance of a course on animal learning to understanding her pet
than she would have in the 1960's.
In the 1980's, the laboratory science of animal cognition
continued to emerge as a discipline in its own right and to
strengthen its conceptual links to the powerful field of human
cognition. There were still more demonstrations of similarities
between animal and human cognition. The idea of attention was
developed further in the context of the phenomena of learned
helplessness and blocking, and began also to appear in new contexts,
including visual search, where perception was sometimes described in
quite human-like terms (see Blough,
2001).
An increasing number of studies appeared on presumably emotional
aspects of animal life, including phobias, aggression, frustration,
reproduction, territoriality, mate selection, bonding, and many
others. Yet, the single most dominant focus was still on an
emotionally detached intellectual capability. We suspect that our
hypothetical student in a class on learning and cognition might
still have been nearly as disappointed in the 1980's with the
relevance of the course to furthering her empathic understanding of
her pet parrot as she would have been in earlier decades.
We feel it may be important at this point to interject that we do
not propose that the success of a course in animal learning and
cognition should necessarily be evaluated in terms of whether
students feel the course improves their understanding of or empathy
for the everyday behavior of their pets. We do predict, however,
that such a course will have a reduced impact on a student’s
subsequent behavior as a citizen if it fails to contact the student’s
intuitive ideas about nonhuman animals. After all, the student’s
intuitive ideas are acquired through everyday life in his or her
culture, so that if there is little or no contact between intuition
and the course, the implication is that there is little contact
between the course and the surrounding culture. We are doubtful that
that kind of intellectual and emotional isolation between a science
and its surrounding culture is beneficial for either.
Is the mental life of animals portrayed differently in the late
1990's? Indications of mental continuity have been demonstrated in
still more ways, including our three specific examples of attention
described above. Perhaps our imaginary student in a class on animal
learning and cognition accordingly might feel a somewhat stronger
link between the course and her feelings and thoughts about her pet
parrot. The beginning student in many American psychology
departments is probably somewhat less likely to be chastised for
talking about memory and attention, so that at least, the concepts
to which she is exposed in the class are more likely to be framed in
a language more conducive to motivating her to think about the
relation between the course and everyday behavior of her pet.
We would summarize the field of animal cognition as still
largely, although certainly not exclusively, focused, however, on
intellectual functioning independent of issues of emotionality,
personality, or social processes. This is despite increasingly many
counter examples. See, for example, De Waal (1999) for a sharp
exception to the general rule. The kind of empathic understanding a
pet owner may be seeking is not prominent in the contemporary
scientific study of the animal mind. The historical indebtedness of
animal cognition to experimental psychology as experimental
epistemology is obvious: The image of an animal that emerges from
such texts and courses mirrors the prototypical image of humans in
contemporary human cognition: It is still that of an
information-processing machine to which feelings and emotions,
especially positive feelings appear relatively seldom (Seligman,
quoted in Ruark, 1999). We acknowledge that some see this as a virtue
(Kendler, quoted in Ruark, 1999)
because they believe it preserves the objectivity of science. We
think, however, that it is problematic that the science, including
its textbooks, scarcely acknowledges the enormous, incalculably
diverse roles played by nonhuman animals in our society. The present
volume on avian cognition exemplifies how little contact the field
of avian cognition has with these non-scientific roles that are
quite central to the normal functioning of our culture. From the
perspective of arousing student interest based on what students
already know and feel about their pets, the field of animal learning
and cognition is still "narrow" by virtue of adhering to a
particular kind of "objectivity," historically associated
with positivism and the machine metaphor for the mind, nonhuman
animal or human.
We ourselves are scarcely immune to the virtues of this
objectification of the animal mind. Our own three examples of
research on avian visual attention reveal their intellectual
heritage in a science that acts as though it is independent of the
surrounding culture, including human values. The science of animal
cognition, like so much of contemporary science, continues to derive
mainly from the belief that science and human values can be, and
quite probably should be, kept separate. The politicization of
animal cognition, we predict, would not be generally welcomed by
most practitioners of the rigorous laboratory science on the animal
mind. Notice the tension that therefore immediately arises.
Scientific practice in animal cognition tends to be based on the
assumption that science should be objective, dispassionate, and
neutral, and therefore should stand apart from everyday human
political society, which involves passionate battles over basic
human values. Yet, scientific practice in animal cognition also
demonstrates dramatic similarities between nonhuman animal and human
minds, so that it becomes more problematic to assume that there is
no set of "animal values" analogous to human values or
that human values pertain only to humans without our having to
examine any of their possible implications for the role of non human
animals in human society. We shall next discuss some difficult
questions this tension poses, the tension, that is, between the
study of the animal mind by a rigorous, dispassionate science, on
the one hand, and the human values of the culture within which that
science is evolving, on the other.
III. Nonhuman Animal Cognition and Human
Values
We showed above that the content of a course on animal learning,
memory, attention and other cognitive processes has changed over the
years from a strictly behavioral approach to a much more mentalistic-sounding
approach. We think such drastic changes are likely to be driven not
just by the internal logic of laboratory research but also by
external, cultural forces. What is the cultural context within which
human scientific understanding of the animal mind is developing? How
does human society portray how laboratory science investigates
animal behavior? Is it possible that human values embedded in human
society affect how the science of the animal mind develops? If so,
can we actually identify any ways in which changing human attitudes
toward this laboratory science might contribute to changes in that
science? We feel these questions need to be addressed. At the
present time, there is scarcely any organized attempt to answer
them. In this section we therefore begin to speculate about these
and other issues, in the hopes that we thereby will promote more
systematic investigation of them.
Consider some examples of how culture might affect the science of
the animal mind.
First, animal facilities and animal care management in the
Psychology Department at the University of Utah are not unique in
being vastly improved over their condition of 30 to 35 years ago.
There is improvement in terms of physical health of the animals and
of reduced health risks of those who work with the animals. There is
also improved oversight in terms of daily animal care, justification
for proposed research, ensuring appropriate euthanasia methods, and
so on. Our facilities have greatly benefitted from the Institutional
Animal Care and Use Committee, the American Association for
Accreditation of Laboratory Animal Care (AAALAC) accreditation,
consequences of the Animal Welfare Act, and so on. It would be
vainglorious and inappropriate self congratulatory behavior on our
part to claim that the improvements we have experienced in our
facilities are due purely to our own activities. It is clear that
the public’s interest in animal welfare has partly facilitated
bringing about these improvements in the housing and care of animals
used in laboratory research on the animal mind. This example
suggests a link between culture and the infrastructure of science.
If there is a link at the level of infrastructure, why could there
not be a link at the direct level of theories of the nonhuman animal
mind?
Second, consider that in the popular press,
"conditioning" is often viewed as dehumanizing.
Conditioning was originally accepted in the 1920's and 1930's by
many hopeful members of the public who, thanks in part to
proselytizing popular articles by Watson and others, saw in
conditioning an egalitarian perspective in terms of which a human
democratic society could be dramatically improved simply by changing
the environmental context within which children developed and in
which adults worked and played. Skinner (1971) enlarged on this
optimistic view in his book, Beyond Freedom and Dignity. More
recently, however, conditioning has perhaps more often been
associated in the public mind with Orwell’s 1984, Burgess’s
Clockwork Orange, countless science fiction books and movies
involving brain washing, and the mechanical, automaton view of the
human condition in terms of which there is no free will and in that
sense no control over one’s life by one’s inner mental life. Novels
and movies portray evil or misguided scientists controlling behavior
by means of conditioning methods. When machine and human values
collide, the cultural results are often appalling, as in the diverse
cases of Mary Shelley’s Frankenstein, Stevenson’s The
Strange Case of Dr. Jekyll and Mr. Hyde, and in the recent novel
by Bakis, Lives of the Monster Dogs. Is it not possible that
this negative popular image of conditioning has contributed in many
different ways to the reduced centrality of conditioning in
laboratory science on the animal mind? We certainly would not want
to argue that the public has an entirely negative view of
conditioning, for such is certainly not the case. Indeed, the public
no doubt has a favorable view of some aspects of applied behavior
analysis by virtue of successful behavioral treatment programs based
on behavior therapy, token economies, and other methods to treat
drug addiction, alcoholism, and various behavior disorders. However,
it does seem to us that the undeniable loss of centrality of
conditioning in the scientific study of animal intelligence may to
some degree derive from pressures emanating from the public.
Third, while popular culture in some ways may take a dim view of
conditioning, it also sometimes takes a very favorable view of
animal intelligence, broadly defined. The public seems to take
delight in both anecdotes and scientific studies suggesting new and
unsuspected forms of animal intelligence, especially when
accompanied by the possibility of some sort of feeling of empathic
understanding. Some of the most beloved American literature of the
last 100 years has involved sentient and verbal nonhuman animals,
such as in E. B. White’s Charlotte’s Web, Charles Schulz’s
Peanuts, Albert Bigelow Paine’s The Hollow Tree
Snowed-in Book, etc., etc. The public is exposed to a constant
barrage of human interest stories about animals in newspapers, on
radio and television, and the number of programs devoted to
naturalistic behavior of animals is correspondingly enormous. We
would not be at all surprised if the public’s interest has
contributed to the rise of the scientific field of comparative
cognition. The field is still to this moment, however, strikingly
independent from the enormous, incalculably diverse roles played by
nonhuman animals in human society. The present volume on avian
cognition exemplifies how little contact the field of avian
cognition has with these non-scientific roles that are quite central
to the normal functioning of our culture. The emotional issues so
prominent in popular culture addressing nonhuman animals, as in the
charming recent book on the emotional role of dogs in human society,
A Pack of Two (Knapp, 1998) and in many other related books
such as When Elephants Weep (Masson and McCarthy, 1995), The
Human Nature of Birds (Barber, 1994), The Minds of Birds,
(Skutch, 1996), Diary of a Pigeon Watcher (Schwerin, 1976), Next
of Kin (Fouts, 1997), Adam’s Task: Calling Animals by Name,
(Hearne, 1987), Why We Love the Dogs We Do: How to Find the Dog
that Matches Your Personality (Coren, 1998), Great Apes,
(Self, 1997), Timbuktu (Auster, 1999), The Animal Estate
(Ritvo, 1987) and Inside the Animal Mind (Page, 1999).
Countlessly many shorter pieces address these issues. One of
shortest and best is by E. B. White (1940/1966) on dog training.
Little contact as the field of comparative cognition has with this
virtually boundless literature on the emotion lives of animals, we
feel comparative cognition makes stronger and more immediate contact
with it than does the literature on a remorselessly
"objective" form of conditioning. The public’s interest
in animal intelligence may therefore promote a cognitive approach to
the animal mind more than a conditioning approach, especially in
view of a perhaps somewhat negative view of conditioning in general.
Fourth, consider a few ways nonhuman animals contribute to human
welfare. A) Animals play a role in humans’ everyday emotional
lives. Animals are increasingly seen as providers of emotional
support for the aged and for both children and adults in hospitals.
In terms of human safety and social integration, guide dogs for the
blind are widely respected and admired. It is hard to see Descartes’
purely mechanical attributions about animals at work in situations
like these, where emotional intimacy and attachment between animal
and human implies that many people believe animals, dogs especially,
can understand how we feel. Admittedly, dogs are a special case
because of their social structure and pair bonding, but the general
possibility that animals have rich emotional lives that could, and
sometimes do, facilitate our own, is clear. B) The use of animals in
biomedical research, while controversial, remains for many a
literally life-giving role. C) Even more controversial is that
animals contribute to human society by providing food and clothing.
Note that this contribution seems at sharp odds with the
contribution to human welfare through bonding and other positive
emotional dimensions. We believe it is one of the as yet unresolved
contradictions in our society that we see nonhuman animals in
impersonal and unfeeling ways that permit us to ethically justify
many of the ways in which we use animals for our own benefit, and
simultaneously, we see animals in sufficiently personal ways to
permit them to provide powerful emotional support. D) Nonhuman
animals in American society contribute to public welfare through
entertainment in movies, circuses, and so on.
Fifth, one of the strongest determinants of how we perceive
nonhuman animals is probably to be found in our religious beliefs.
An illuminating example is provided by a recent volume (Larson,
1997) on the 1925 Scopes trial in Dayton Tennessee, in which a high
school biology teacher was tried and convicted for breaking a
Tennessee law banning the teaching of Darwinian ideas about human
evolution. The author, E. J. Larson, provides a wonderfully
interesting account of how fundamentalist Christian beliefs and
attitudes shaped Tennessee state law on teaching about the relation
between human and nonhuman animals. It is worth noting in passing
that different cultures view this relation in dramatically different
ways. A cross-cultural analysis could be conducted either in
historical terms or in terms of different contemporary cultures. We
expect an exploration of ancient Egyptian, Native American, Hindu,
Buddhist, as well as Christian and Jewish views would illuminate
radically different and therefore highly interesting cultural
differences in attitudes and beliefs about animals. It would be
interesting to investigate whether different religious beliefs have
contributed to differences across cultures in the development of a
laboratory science of the animal mind.
Sixth, consider the discussion, pro and con, on the political,
ethical, economic, and philosophical movement for animal rights,
animal welfare, and animal law (e.g., Orlans, Beauchamp, Dresser,
Morton, and Gluck, 1998; Petrinovich, 1999; Preece, 1999; Preece and
Chamberlain, 1993; Regan and Singer, 1989; Singer, 1990). This
movement may have facilitated the improvement of animal care in
animal shelters (Tuber, Miller, Caris, Halter, Linden, and Hennessy,
1999) as well as in research laboratories, and the movement’s
political goals can be expected to have further implications for
scientific research involving nonhuman animals (Plous, 1998). It is
hard to imagine a researcher studying the animal mind who has not
already been affected by this movement and all signs point to
greater impact in the future.
Seventh, our culture is famous for its rapid growth in
technology, especially as relating to computers. It seems inevitable
to us that the technology of the surrounding culture affects the
kinds of experiments researchers choose to conduct with nonhuman
animals. For example, of the three experiments on attention we
described earlier, we could have conducted only one, that on spatial
attention, with technology generally available to the behavioral
scientist before roughly 1980. We think it is reasonably clear that
nonhuman animal research using complex visual stimuli and reaction
times could not have been conducted before the arrival of
inexpensive laboratory computers. Since complex stimuli having
specified quantitative properties are essential to much research on
avian visual cognition, this is equivalent to saying that avian
cognition is highly dependent on the surrounding culture,
specifically, on the culture’s technological resources. This is
but one of many examples one could give of how research in one
domain depends on techniques and ideas developed elsewhere in a
culture.
In summary, human society’s beliefs about animals are
incredibly diverse, and seem to offer ample opportunity to encourage
some approaches to the scientific study of the animal mind and to
discourage others. Human society is in many ways sharply self
contradictory in its positions on nonhuman animals, with some
beliefs compatible with a Cartesian machine-like and value free
view, and some human-like and value laden. We suggest that a more
careful and in-depth historical and cultural analysis of these
beliefs may contribute to an understanding of why the scientific
study of animal intelligence changes the way it does.
Integrating Science and Human Values
in the Psychology Classroom: A Service-Learning Course
We have seen that there are increasingly many demonstrable
similarities between nonhuman animal and human minds, that the
scientific study of the animal mind tends to focus on dispassionate,
intellectual characteristics, that there seems to be a historical
trend away from conditioning and toward cognitive accounts of the
animal mind, that human society makes use of animals for, among many
other things, highly personal emotional needs, and that there are
many powerful cultural beliefs that might affect the development of
a science of the animal mind. Let us now examine a method we have
begun to use to study interrelations among these diverse scientific
traditions and cultural forces. Our method is a Service-Learning
course on comparative cognition offered at the University of Utah.
The course is called "The Animal Mind in Nature, the
Laboratory, and Human Culture."
We wish to caution the reader against the optimistic but we fear
unrealistic hope of developing a coherent picture of these
interrelations. We merely wish at present to describe a way in which
to begin to discuss them in an academic setting. At present, we
believe we will need to accept that so many of these relations are
in such stark opposition to one another that an attractively
coherent account may not be possible, at least not for the
foreseeable future.
Our class has two mutually defining components. The first is the
usual academic component of a class in animal learning and
cognition. In this part of the class, which consumes nearly
two-thirds of a semester term, a student learns basic methods,
facts, and theories, with a relatively strong dose of intellectual
history, including 17th to 20th century
philosophy of mind. We sometimes try to illuminate the diverse
scientific perspectives by contrasting behavioral and cognitive
accounts. We usually use the computer simulation technique as
embodied in Catania and Shimoff’s Behavior On a Disk
to illustrate operant conditioning phenomena, and we discuss journal
articles on laboratory studies of short-term memory, spatial memory,
visual discrimination, the three types of attention described above
in sections II - IV, problem solving, transposition, and choice
behavior, and naturalistic behaviors such as navigation and caching.
The second component of our class is the service-learning
component. Students in our course participate not only in
traditional academic lecture and discussion sessions, but also
integrate the traditional course content with real world experiences
as volunteers at community organizations directly involved with
nonhuman animals. Students volunteer three hours per week at a
placement they choose from a list we provide. Volunteer activities
have involved several different types of activities. Some activities
are designed to facilitate the development and implementation of
"behavioral enrichment" methods at the local aviary and
zoo. Some students, for example, are assigned the task of inventing
and constructing novel devices and "toys" to encourage
more naturalistic behaviors by birds in captivity at the Tracy
Aviary. Another category of activity involves collecting
observational data to facilitate the development of ethograms to
show how animals in captivity spend their time engaged in different
behaviors in different locations in their cages. Students have
recorded observational data from gorillas and chimpanzees. Students
particularly empathized with Gorgeous, a lowland gorilla who
recently died of causes due to extreme old age. When Gorgeous and a
student looked into each other’s eyes, a student acquired a whole
new meaning of animal cognition. The observational data collected at
the zoo will be used in the design of improved facilities now in the
planning stages. A third category of activity involves serving as a
"greeter" at the Salt Lake County Animal Shelter, where
members of the public need to be instructed in the policies and
functions of the Shelter, illegal exchanges of animals in the
shelter parking lot need to be prevented, and grieving pet owners
searching for their lost pets need to be consoled and informed about
shelter policies. The shelter’s chief goal is to educate the
public on the issue of responsible pet ownership, and students
volunteering at this placement learn a good deal about public policy
and ethical issues involved in pet ownership, and they directly
share both traumatic and joyful experiences with pet owners.
Students observe and empathically experience the despair when pets
are not at the shelter when owners come to look for them. Students
who describe such experiences in class may be so overwhelmed that
they cry at the memory of the joy or despair experienced by both the
owner, and apparently, an animal pet. The "animal mind" is
not a dispassionate and purely intellectual issue to a student who
has shared such personal experiences. Still other volunteer
placements have involved students’ helping to train dogs at the
Salt Lake Dog Training Club. Students who do not wish to directly
interact with animals have had the opportunity to become involved
with the State Division of Wildlife Resources, where students
observe meetings held to determine public sentiment about possible
new state wildlife policy and to educate the public on new policies.
Students consult with Utah State staff to suggest ways to coordinate
state wildlife management policy with the public’s wishes.
Students observe the public’s considerably emotional reactions at
some of these public meetings held to establish or review public
policy on hunting and on the return of injured or illegally trapped
animals to the wild. Students clearly see in such cases how public
policy on nonhuman animals affects the everyday lives of members of
the community in ethical, economic, and political ways.
Students in each placement setting see academic course material
from lectures and readings applied in the real world, and try to
interrelate academic course content with broader cultural issues. In
addition to becoming familiar with how laboratory science views
animals in a relatively detached, unemotional way, students
empathically experience many of the community’s diverse
interactions with nonhuman animals. We feel that one of the most
valuable consequences of students’ efforts to apply laboratory
material to their service-learning experiences is their better
appreciation of the difference between scientific and everyday
approaches to nonhuman animal behavior. Students acquire sharper
awareness of critical implications of scientific skepticism for the
attributions they customarily make about knowledge, perceptions, and
"awareness," in nonhuman animals. They acquire a new
sensitivity to the scientifically problematic nature of their easy
attributions about their pets. And, they acquire a new sensitivity
to the difficulty of knowing whether any of their usual beliefs
about their pets are scientifically correct, or even meaningful.
That is, the juxtaposition of scientific and everyday accounts of
nonhuman animal behavior sharpens a student’s awareness of the
difference between them. We feel this sharpened awareness
facilitates a student’s appreciation of the corresponding
difference between scientific accounts of human behavior and
"pop psychology" accounts.
We try to ensure that each student learns about the
service-learning experiences of other students. On a regular basis,
class meetings are set aside for group discussion of student
placement activities. Students keep journals in which they describe
their volunteer service; each week they write a journal page
describing their experiences in the past week, reflect on those
experiences, and interrelate them to the academic objectives of the
course. Each student also writes a paper due at the end of the
academic term which addresses some issue of deep personal concern to
the student. The main criterion for the selection of a topic is that
it address some issue involving nonhuman animals a student finds to
be deeply involving, emotionally uplifting or disturbing, or some
issue that confronts a deeply held belief. Topics therefore vary
widely, and include ethical issues involved in the use of nonhuman
animals in laboratory research, including issues deriving from
genetics research and cloning, ostensible occurrences of attentional
phenomena in animals observed at the zoo, limitations on what one
can know what another person or an animal knows or feels, possible
evolutionary explanations for why family pets do or do not
emotionally bond with family members, the emotional and
psychological needs met by hunting, the relation between animal
abuse and spousal abuse, ethical issues involved in keeping animals
in caged environments, traditional Native American compared to
Christian views of nonhuman animals, and many others. We feel that
these student papers are successfully sensitizing students to
interrelations between their own personal values and laboratory
research on the nature of animal intelligence.
In short, students are encouraged to personally experience
culturally pervasive but often individually unexamined relations
between animals and humans, and to consider the reciprocal
implications of cultural beliefs for the laboratory science of the
animal mind and of the science for cultural beliefs. Students seem
generally to agree that these experiences bring the academic subject
matter of comparative cognition alive and make the personal,
emotional aspects of the roles of animals in human society clearer.
The value-laden nature of some of these roles becomes much clearer
when students personally experience them. It becomes clear that
personal religious positions are critical to how students react to
these roles. Generally, students begin to seriously consider that
what they perceive an animal to be depends in part on culture and on
intensely held personal beliefs, as well as on what contemporary
science says an animal is by virtue of laboratory results on
attention, memory, and learning. While a student’s intellectual
growth along these lines makes comparative cognition appear much
more complex, it also makes it appear much more interesting and more
relevant to the student’s own life.
The Structure of this Chapter Exemplifies the
Tension Resulting from Polarized Views on the Animal Mind
There is a central tension in the "culture wars" or
"science wars" between the sciences and the humanities.
This tension is so severe at times that it can wall off entire
academic departments from each other, say Neuroscience from Women’s
Studies. One part of this tension is sometimes focused on the basic
nature of the human condition, with some being partial to an
objective "scientific" view and with others preferring to
make "values" central to the definition of a human being.
We hope we have reminded the reader that a related and equally
strong tension exists between the idea that nonhuman animals are
machines, on the one hand, and the idea that they are creatures
supporting human empathy, on the other. This tension is addressed by
a growing scientific and philosophical literature from which a
satisfactory means of resolving it may emerge in the future (Allen
and Bekoff, 1997; Petrinovich, 1999; Preece, 1999). Today, however,
there is no such way, and therefore this chapter is essentially two
inadequately connected chapters, with the inadequacy reflecting how
our culture fails to integrate the two perspectives on what a
nonhuman animal is. First, we describe laboratory research on human
and avian attention. This research typifies a common
"value-free" approach according to which science is
objective, impersonal, and is seen as relatively independent of the
morass of conflicting human values in the surrounding culture.
Second, we describe how people feel about nonhuman animals,
especially pets, and we contrast this subjective and personal view
with the scientific view. Our service-learning course addresses and
clarifies the extreme tension between these two views, but we are
acutely aware that there are no tools available, either for the
reader or for the authors, by which the two views can be reconciled
and the tension released. We offer no way to synthesize these
divergent views. This chapter reflects these sharply polarized views
and of necessity it too displays unresolved tensions and
contradictions. We hope a reader will be able to accept this tension
and to be able to profit from it in the sense of having a clearer
appreciation of how science and culture offer contrasting and, we
trust, mutually informing insights on nonhuman animals. We hasten to
note that we make no claims to being the first to identify this
tension: Many readers happily familiar with the Far Side cartoons
by Gary Larson will have already experienced it.
A Prediction
The scientific study of animal intelligence, including visual
attention, typically makes an implicit distinction between
intellectual and emotional components of an animal’s life, in the
sense that the science usually acts as though its proper goal is a
purely objective analysis of the nonhuman animal mind. When such a
scientific goal is framed within Western culture, it implies
emotions, vague feelings, and other quintessentially subjective
phenomena should be excluded, because otherwise they are likely to
cloud an otherwise pristine and objective portrait of animal
intelligence. Furthermore, the science most often views itself as
best separated from human values as they appear in the highly
emotional world of everyday politics. We admit this characterization
oversimplifies the science because in fact much biobehavioral
research attempts to integrate intellectual functioning in learning
and memory with processes that surely involve emotional features,
such as reward, fear, mate selection, and many other functions, and
researchers and professional organizations of psychologists are
increasingly calling attention to the value of nonhuman animal
research for general human welfare. Nevertheless, we feel that on
the whole, the science prefers to treat animal intelligence as
sharply distinct from intrinsically subjective processes. In short,
the science often implicitly assumes that animal intelligence can be
studied apart from any nonhuman animal emotional life. The
corresponding assumption in the study of the human mind is highly
controversial: human intelligence is increasingly seen not as
something which can be studied apart from subjective features of
human life but instead as dependent on subjective elements of human
culture, including human values. A corresponding position on the
nonhuman animal mind would acknowledge that animal intelligence
depends in some way on subjective elements. We expect this latter
position to become more prominent because research continues to
encourage the position that human and non human cognitive processes
have much in common.
It is not a commonly discussed possibility in the contemporary
science of the animal mind, at least not the avian mind, that the
metaphors, analogies, and plain English phrases used in that science
depend on extra scientific culture for their meanings. Hacking
(1999) describes several examples of this dependency in the social
and behavioral sciences. One in particular is applicable here and
highlights both the point we wish to make regarding avian cognition
and the kinds of potential pitfalls that rightly worry rigorous
scientists and scholars interested broadly in philosophy of mind.
Hacking (1999, p. 64) writes that "We are not surprised to hear
that the results of primatology bear strong traces of their
discoverers. We can well imagine what Donna Haraway (1989) and
others have taught us in detail: accounts of the behavior of
primates reflect the societies of the scientists who study them. We
all know the bad jokes about British apes with stiff upper lips,
ruthlessly enterprising American apes, hierarchical and
communitarian Japanese apes, promiscuous French apes. Primates,
perhaps, have been a field for working out ourselves as much as
describing animal communities." We take away two messages here.
First, it would not be surprising that as the study of the avian
mind progresses, and in particular as increasingly many similarities
are discovered between the avian mind and the human mind, the
picture that emerges of the avian mind will resemble in part the
culture of the scientists constructing that picture. Second, as a
result, it will be increasingly important for scientists responsible
for this picture to scrutinize it carefully for any features of
themselves. This scrutiny will be necessary to uncover what we
suspect will be the inevitable unexamined assumptions inadvertently
left in the picture as a residue of the culture in which the
scientists work. We are not at all certain that the scrutiny will in
fact reveal all these residual assumptions, but the effort will
contribute to a better understanding of the relation between the
human cultures in which behavioral scientists work and emerging
scientific pictures of the avian mind. We therefore feel the science
of the animal mind can benefit in several ways from a more
systematic examination of its dependence on surrounding cultural
beliefs, considering the obviously vast economic, political, and
ethical values, beliefs, and assumptions about nonhuman animals in
our culture.
In summary, we anticipate the future will see a more careful
scrutiny of how cultural beliefs about the lives of nonhuman animals
affect the development of a science of the nonhuman animal mind, and
more generally, of what we might call the "animal
condition." We anticipate that this more careful scrutiny will
facilitate determining how the mental lives of nonhuman animals and
of humans are similar or different. We are inclined to believe that
human culture is a pervasive and powerful determinant of how even
scientists trying very hard to work independently of it view the
nonhuman animal mind . We predict culture will therefore continue to
influence how scientists conceptualize and prioritize the problems
they study. In the short term, we may see, for better or worse, a
continuing shift of the science of the nonhuman animal mind,
including the avian mind, in a direction compatible with the
surrounding culture’s view of the relation between humans and non
humans. At the same time, it is to be hoped that culture’s view
will continue to be influenced by the science of the nonhuman animal
mind. We hope in this chapter to have called attention to the value
of asking what will be the resulting interaction between science and
culture.
IV.
References
Allen, C., & Bekoff, M. (1997).
Species of mind. Cambridge, MA: The
MIT Press.
Ashby, F. G., & Gott, R. E. (1988). Decision rules in the
perception and categorization of multidimensional stimuli. Journal of
Experimental Psychology: Learning, Memory, and Cognition, 14, 33-53.
Ashby, F. G., & Maddox, W. T. (1998). Stimulus categorization. In
M. H. Birnbaum (Ed.), Measurement, judgment, and decision making:
Handbook of perception and cognition (pp. 251-301). San Diego, CA:
Academic Press.
Auster, P. (1999). Timbuktu. New
York, NY: Henry Holt.
Bakis, K. (1997). Lives of the
monster dogs. New
York, NY: Warner
Books.
Barber, T. X. (1994). The human
nature of birds. New
York, NY:
Penguin.
Blough, D. S. (2001).
The perception of similarity. In R. G. Cook, (Ed.),
Avian visual cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/dblough/
Branch, M. N. (1977). On the role of "memory" in the analysis
of behavior. Journal of the Experimental Analysis of Behavior, 28,
171-179.
Budiansky, S. (1998). If a lion
could talk: Animal intelligence and
the evolution of consciousness. New York, NY: The Free Press.
Burgess, A. (1962).
A Clockwork orange. New
York, NY: Norton.
Bushnell, P. J. (1995). Overt orienting
in the rat: Parametric studies of cued
detection of visual targets. Behavioral
Neuroscience, 109, 1095-1105.
Candland, D. K. (1993). Feral children & clever animals:
Reflections on human nature. Oxford: Oxford University Press.
Catania, A. C., Matthews, B. A., &
Shimoff, E. (1997). Behavior on a Disk (Version 4.0) [Computer software]. Columbia, MD: CMS Software.
Chase, S. &
Heinemann, E. G. (2001). Exemplar memory and discrimination. In R. G.
Cook, (Ed.), Avian visual
cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/chase/
Clayton, N. S., & Dickinson, A. (1998). Episodic-like memory during cache
recovery by scrub jays. Nature, 395, 272-274.
Clayton, N. S., & Dickinson, A. (1999). Memory for the content of caches by
scrub jays (Aphelocoma coerulesceus). Journal of Experimental Psychology:
Animal Behavior Processes, 25, 82-91.
Cook, R. G. (2001). Hierarchical
stimulus processing in pigeons. In R. G. Cook, (Ed.),
Avian visual cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/cook/
Coren, S. (1998). Why we love the dogs
we do: How to find the dog
that matches your personality. New York, NY: The Free Press.
De Waal, F. B. M. (1999). The pitfalls of not knowing the whole animal.
The Chronicle of Higher Education, XLV, B4-B6.
Douglas, M., & Hull, D. (Eds.). (1992).
How classification works: Nelson
Goodman among the Social Sciences. Edinburgh, U.K.: Edinburgh University Press.
Fouts, R. (1997). Next of kin. New
York, NY: Morrow.
Fremouw, T., Herbranson, W. T., & Shimp, C. P. (1998). Priming of
attention to local or global levels of visual analysis. Journal of
Experimental Psychology: Animal Behavior Processes, 24, 278-290.
Goodman, N. (1977). The structure of
appearance. Boston, MA: K. Reidel.
Hacking, I. (1999). The social construction of
what? Cambridge, MA:
Harvard University Press.
Hanson, N. R. (1969). Perception and
discovery. San Francisco, CA:
Freeman, Cooper.
Haraway, D. (1989). Primate visions: Gender,
race and nature in the world
of modern science. London, U. K.: Verso.
Hearne, V. (1987). Adam’s task: Calling animals by name. New
York: Knopf.
Herbranson, W. T., Fremouw, T., & Shimp, C. P. (1999). The
randomization procedure in the study of categorization of multidimensional
stimuli by pigeons. Journal of Experimental Psychology: Animal Behavior
Processes, 25, 113-135.
Huber, L. (2001). Visual
categorization in pigeons. In R. G. Cook, (Ed.),
Avian visual cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/huber/
Inman, A., & Shettleworth, S. J. (1999). Detecting metamemory in nonverbal
subjects: A test with pigeons. Journal of Experimental Psychology: Animal
Behavior Processes, 25, 389-395.
Kagan, J. (1998). Three seductive
ideas. Cambridge, MA: Harvard
University Press
Kamin, L. (1969). Predictability, surprise, attention, and
conditioning. In B. A. Campbell & R. M. Church (Eds.), Punishment.
New York, NY: Appleton-Century-Crofts.
Knapp, C. (1998). Pack of two: The intricate bond between people and
dogs. New York, NY: The Dial Press.
Krechevsky, I. (1932). "Hypotheses" in rats. Psychological
Review, 39, 516-532.
Lamb, M. R., & Robertson, L. C. (1990).
The effect of visual angle
on global and local reaction times depends on the set of visual angles
presented. Perception & Psychophysics, 47, 489-496.
Larson, E. J. (1997). Summer for the
gods: The scopes trial and america’s continuing debate over science and religion. New York, NY: Basic Books.
Masson, J. M., & McCarthy, S. (1995). When elephants weep.
New York, NY: Delta.
Orlans, F. B., Beauchamp, T. L., Dresser, R., Morton, D. B.,
& Gluck, J. P.
(1998). The human use of animals: Case studies in ethical choice. New
York, NY: Oxford.
Orwell, G. (1949). 1984. New
York, NY: Harcourt
Brace.
Paine, A. B. (1910). The hollow tree
snowed-in book. New
York, NY: Harper
and Brothers.
Page, G. (1999). Inside the animal mind. New
York, NY: Doubleday.
Petrinovich, L. (1999). Darwinian dominion: Animal
welfare and human interests. Cambridge, MA: The MIT Press.
Plous, S. (1998). Signs of change within the animal rights movement:
results from a follow-up survey of activists. Journal of Comparative
Psychology, 112, 48-54.
Posner, M. I. (1980). Orienting of
attention. Quarterly Journal of
Experimental Psychology, 32A, 3-25.
Posner, M. I., Nissen, M. J., & Ogden, W. C. (1978). Attended and
unattended processing modes: The role of set for spatial location. In H. L. Pick
& I. J. Saltzman (Eds.), Modes of perceiving and processing information.
Hillsdale, N.J.: Lawrence Erlbaum.
Preece, R. (1999). Animals and nature: Cultural
myths, cultural realities.
Vancouver, BC: University of British Columbia Press.
Preece, R., & Chamberlain, L. (1993).
Animal welfare and human values. Waterloo,
U. K.:
Wilfrid Laurier.
Regan, T., & Singer, P. (1989). Animal
rights and human obligations. Englewood
Cliffs, NJ: Prentice Hall.
Reynolds, G. S. (1961). Attention in the pigeon. Journal of the
Experimental Analysis of Behavior, 4, 203-208.
Riley, D. A., & Roitblat, H. L. (1978). Selective attention and
related cognitive processes in pigeons. In S. H. Hulse, H. Fowler, &
W. K. Honig, (Eds.), Cognitive processes in animal behavior (pp.
249-276). Hillsdale, NJ: Lawrence Erlbaum.
Ristau, C. A. (1991). Cognitive
ethology: The minds of other animals. Hillsdale, NJ: Lawrence Erlbaum.
Ritvo, H. (1987). The animal estate: The
english and other creatures
in the victorian age. Cambridge, MA: Harvard.
Robertson, L. C. (1996). Attentional persistence for features of
hierarchical patterns. Journal of Experimental Psychology: General, 125,
227-249.
Romanes, C. J. (1898). Mental evolution in animals. New
York, NY:
Appleton. (Original work published 1883)
Rorty, R. (1980). Philosophy and the
mirror of nature.
Princeton, NJ: Princeton.
Rosch, E., & Mervis, C. B. (1975). Family resemblance: Studies in
the internal structure of categories. Cognitive Psychology, 7,
573-605.
Ruark, J. K. (1999, February 12). Redefining the Good Life; a
New Focus in the Social Sciences. Chronicle of Higher Education
(p A14).
Schulz, C. M. (1968)."He’s your dog, Charlie Brown!" Cleveland,
OH:
World Publishing Co.
Schulz, C. M. (1971). Snoopy and "It was a dark and stormy
night." New York, NY: Holt, Rinehart & Winston.
Schwerin, D. (1976). Diary of a
pigeon watcher. New
York, NY:
Paragon House.
Self, W. (1997). Great apes. New
York, NY: Grove.
Shelley, M. W. (1969). Frankenstein, or the modern
prometheus.
London, U. K.: Oxford. (Original work published 1818)
Shimp, C. P. (1976). Organization in memory and
behavior. Journal of
the Experimental Analysis of Behavior, 26, 113-130.
Shimp, C. P. (1982). Metaknowledge in the pigeon: An organism’s
knowledge about its own behavior. Animal Learning and Behavior, 10,
358-364.
Shimp, C. P. (1983). The local organization of behavior: Dissociations
between a pigeon’s behavior and self-reports of that behavior.
Journal of the Experimental Analysis of Behavior, 39, 61-68.
Shimp, C. P., Fremouw, T., Ingebritsen, L. M., & Long, K. A. (1994).
Molar function depends on molecular structure of behavior. Journal of
Experimental Psychology: Animal Behavior Processes, 20, 96-107.
Shimp, C. P. & Friedrich, F. J. (1993). Behavioral and computational
models of spatial attention. Journal of Experimental Psychology: Animal
Behavior Processes 19, 26-37.
Shimp, C. P. (1999). Tolerance in a Rigorous
Science. Journal of
the Experimental Analysis of Behavior, 71, 284-288.
Singer, P. (1990). Animal liberation (2nd ed.). New York, NY: Random House.
Skinner, B. F. (1971). Beyond
freedom and dignity. New
York, NY:
Knopf.
Skinner, B. F. (1989). The origins of cognitive thought. American
Psychologist, 44, 13-18.
Skutch, A. F. (1996). The minds of
birds. College
Station, TX: Texas
A & M.
Smith, J. D., Shields, W. E., Allendoerfer, K. R., & Washburn, D.
A. (1998). Memory monitoring by animals and
humans. Journal of
Experimental Psychology, 127, 227-250.
Stevenson, R. L. (1907). The strange case of Dr. Jekyll and Mr. Hyde.
New York, NY: Scribners. (Original work published 1886)
Sutton, J. E., & Roberts,
W. A. (2001). Attentional processes in compound stimulus processing by
pigeons. In R. G. Cook, (Ed.),
Avian visual cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/sutton/
Tinbergen, L. (1960). The natural control of insects in pinewoods: I.
Factors influencing the intensity of predation by songbirds. Archives
Neerlandaises de Zoologie, 13, 265-343.
Tolman, E. C. (1932). Purposive behavior in animals and men. New
York, NY: Appleton-Century-Crofts.
Tuber, D. S., Miller, D. D., Caris, K. A., Halter, R., Linden, F.,
& Hennessy, M. B. (1999). Dogs in animal shelters: Problems, Suggestions, and
Needed Expertise. Psychological Science, 10, 379-386.
Urcuioli, P. (2001).
Categorization & acquired equivalence. In R. G. Cook, (Ed.),
Avian visual cognition [On-line]. Available: pigeon.psy.tufts.edu/avc/urcuioli/
White, E. B. (1952) Charlotte’s
web. New
York, NY: Harper and Row.
White, E. B. (1966). Dog Training. In One
man’s meat. New
York, NY:
Harper & Row. (Original work published 1944)
Wittgenstein, L. (1953). Philosophical investigations. New
York, NY:
Macmillan.
|
Acknowledgment
Some of the empirical research described in this article was supported
in part by grants from NSF and NIMH. We would like to thank Linda Bonar,
Renee Buchanan, and Irene Fisher of the Lowell Bennion Community Service
Center at the University of Utah, and also the Psychology Department of
the University of Utah, for support of the development and implementation
of the Service-Learning course, Psychology 3110, The Animal Mind in
Science, Nature, and Human Culture. We would also like to thank Sarah
Grison and Heather Hartman for their help as graduate teaching assistants
for the course, and Kristen Kamerath and Layne Goble as undergraduate
Bennion Center Teaching Fellows. Special thanks are due the many
service-learning placement coordinators and staff, without whose help the
course could not have been developed. Last but not least, special thanks
are due to the undergraduate students in Psychology 3110, whose many
insights and passionately held beliefs have greatly facilitated the
development of this course.
Correspondence regarding this article should be sent to: Charles P.
Shimp, Department of Psychology, University of Utah, 390 S. 1530 E. RM 502
Salt Lake City, Utah 84112-0251, Telephone: 801-581-8483. Electronic
mail should be sent to: Shimp@psych.utah.edu
Fax:801-581-5841
|
|
