Animals both in nature and in the laboratory often have to
respond to multidimensional stimuli. For instance, animals in nature
typically forage through complex environments where food items must be
recognized against various backgrounds. The successful detection
of target food items depends on the ability to filter out non-food background
items. Furthermore, the animal may need to distinguish between toxic and
safe foods that are highly similar in appearance. Therefore, although a
stimulus may have a number of properties, such as its color, size, shape,
and spatial position, only one of these properties may be relevant for
the problem the organism needs to solve. An animal's ability to respond
to the relevant property is the outcome of attention. If it can isolate
the relevant property and respond only to it, the animal is said to attend
to the relevant dimension. On the other hand, if it responds to other
properties of the stimulus as much as it does to the critical property,
it shows detrimental effects of divided attention and an inability to devote
sufficient attention to the relevant dimension. The degree to which
animals can successfully divide attentional resources is limited by a number
of factors, including the sensory capabilities of the species (see Dukas,
1998 for a discussion of constraints on information processing).
II.
Historical
Antecedents in the Study of Attention
Discrimination
Learning Theories
The process of attention played a key role in an early debate about
the nature of discrimination learning in animals (see review by Riley &
Leith, 1976). In discrimination learning, an animal is presented
with two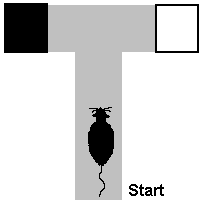 or more stimuli and must learn to respond in different ways to
these stimuli. For example, a rat might have to repeatedly choose
between black and white
goal boxes in a T-maze, with food placed only in the white goal box.
Since the positions of the black and white goal boxes change from left
to right randomly from one trial to the next, the rat must learn to attend
to the brightness of the boxes to find the food. When rats are trained
on this problem, it is often observed that they initially turn repeatedly
to the right or to the left, a behavior called a position habit.
While in a position habit, the rat fails to discriminate between black
and white above a chance level of accuracy. The rat eventually breaks
out of the position habit and progressively shows a preference for the
food-baited white box over the non-baited black box.
or more stimuli and must learn to respond in different ways to
these stimuli. For example, a rat might have to repeatedly choose
between black and white
goal boxes in a T-maze, with food placed only in the white goal box.
Since the positions of the black and white goal boxes change from left
to right randomly from one trial to the next, the rat must learn to attend
to the brightness of the boxes to find the food. When rats are trained
on this problem, it is often observed that they initially turn repeatedly
to the right or to the left, a behavior called a position habit.
While in a position habit, the rat fails to discriminate between black
and white above a chance level of accuracy. The rat eventually breaks
out of the position habit and progressively shows a preference for the
food-baited white box over the non-baited black box.
Debate about the processes involved as rats learned to solve the T-maze
focused on the role of attention in learning. Some theorists proposed that
rats gradually learned to solve the maze through experiences with reinforced
responses over a number of trials. For instance, Spence (1936) proposed
that repeated responses to a stimulus followed by reinforcement (entering
the white box and obtaining food in the example above) led to a tendency
to approach that stimulus (excitation), and repeated responses to a stimulus
that were nonreinforced (entering the black box and finding no food) led
to a tendency to avoid that stimulus (inhibition). These tendencies
were developed over experience with the two choices. Furthermore, all stimuli
impinging on an animal's receptors at the time of response and reinforcement
or nonreinforcement acquired excitation and inhibition, which might include
stimuli other than box brightness or location. The finding that rats initially
displayed position habits in visual discrimination problems was explained
as an effect of pre-experimental differences in the tendency to turn right
or left. Over repeated training trials, the bias was gradually overcome
by the processes of excitation and inhibition that developed to the reinforced
and non-reinforced boxes. Thus, what appeared to be a sudden shift in attention
from position to brightness could be explained as a change in approach
and avoidance tendencies within a continuous learning process.
Theories such as Spence�s that proposed a gradual learning process are
referred to as continuity theories. Other theorists, such as Lashley (1929)
and Krechevsky (1938), argued that the rat attends to only one dimension
of the discrimination problem at a time instead of gradually learning how
to solve the task. Thus, the period of a rat's position habit is
one in which the animal attends only to the dimension of spatial position
(left or right) and not to brightness of the goal box (black or white).
The rat will only learn about the correctness of white over black when
it switches its attention from the position dimension to the brightness
dimension. As Krechevsky (1932) suggested, the rat tried different
hypotheses to solve the task and only learned the correct response when
it hit upon the correct hypothesis. These theorists considered learning
to be noncontinuous and insightful- a distinct shift in attention from
one dimension to another. Such non-continuity theories therefore
assumed that as long as a rat was persisting in a position habit, it was
learning nothing about relative values of black and white.
The numerous studies carried out to test these theories (e.g., Bitterman
& Coate, 1950; Ehrenfreud, 1948; Spence, 1945) led to the conclusion
that animal attention was not as narrow as originally conceived by Lashley
and Krechevsky�s non-continuity theories. Rats appeared to learn
about the differential reinforcement of black and white while responding
only to position. On the other hand, animals did not have unlimited
ability to attend to all stimuli impinging on their receptors, as the continuity
position had proposed.
In an attempt to account for animals� ability to focus attention on
only relevant stimulus dimensions while learning in a gradual way, Sutherland
and Mackintosh (1971) proposed a theory of discrimination learning that
defined neural mechanisms that respond to stimuli along different dimensions
as analyzers. Their theory proposed that discrimination learning
involves two processes, first strengthening the analyzer for the relevant
dimension of the discrimination, and, second, building approach and avoidance
responses to different values or outputs of the analyzer. In the
T-maze example used above, the theory assumes that the brightness analyzer
is gradually being strengthened by consistent reinforcement of white and
nonreinforcement of black. At the same time, the left-right spatial
position analyzer is being weakened by nondifferential reinforcement of
its outputs. The theory clearly argues for differential attention
to stimulus dimensions; because the strengths of all analyzers sum to a
constant value, as one analyzer is strengthened, others must be weakened.
On the other hand, an animal may attend to more than one dimension at the
same time, because two or more analyzers may have sufficiently high strengths
to both control behavior.
Attention
in Information Processing
By the 1960s, a new interest in attention as a factor in information
processing systems had developed. Although this new approach was
concerned with questions about the breadth of attention, as had continuity
and noncontinuity theories of discrimination learning, its focus was otherwise
quite different from these earlier theoretical concerns. Its emphasis
was on attentional processes in humans and not animals. Furthermore,
it stressed innate limits on attention and not the role of attention in
the learning process. Analogies with computers as information processing
systems that had to deal with multiple sources of information simultaneously
were drawn, and limits on attention were conceived of as processing bottlenecks
or overloads.
Human dichotic listening experiments provided a particularly dramatic
demonstration of limited attention. A person was instructed to listen
to a message coming in one ear over a headphone and to shadow or repeat
that message. If information was played into the other ear at the
same time, subjects acquired almost no information about the message presented
to the unattended ear (Cherry, 1953). Broadbent�s (1958) filter theory
was a particularly important account of these findings. He proposed
that a subject attending to a message in one ear selectively blocked or
filtered out the information coming in the other ear. Furthermore,
these early attentional processes were based on the physical properties
of the stimulus and not its semantic content. Later research showed,
however, that a person did pick up some information coming in the unattended
ear based on semantic content. Thus, a person shadowing a message
in one ear would still recognize his own name played in the other ear.
Such discoveries led to the subsequent modification of filter theory.
Treisman (1969), for example, argued that the unattended channel acts as
an attenuator that weakens the unattended message, so that only the most
salient information will reach consciousness.
In other experiments involving more than one sensory modality, subjects
had to attend to one, two, or four dimensions of auditory and visual stimuli
at the same time. In the case of four dimensions, a person could
be asked to judge the pitch or intensity of a tone and the horizontal or
vertical position of a dot on a TV monitor. People's ability to detect
either the visual or auditory stimuli declined as the number of dimensions
processed increased (Lindsay, Taylor, & Forbes, 1968). It was
suggested that the number of dimensional channels a person can process
simultaneously is limited. Thus, detection breaks down as the number
of channels to be processed increases.
III.
Limited
Attention Effects in Pigeons
Soon after researchers began studying attention in human information
processing, an interest developed in investigating attention as a factor
in the information processing systems of pigeons. A
number of different types of experiments have been performed. The
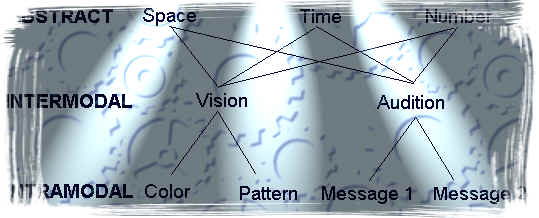
hierarchical framework shown in Figure 1attempts to organize these attentional studies on the basis of the combinations
of sensory input used. At the lowest level of the hierarchy are intramodal
studies, in which a bird must attend to information coming in a single
sensory modality. A compound might consist of two visual elements
within a compound, such as a color and a shape, or two auditory stimuli
of different frequencies presented together. At the next level in
the hierarchy, the intermodal level, compound test stimuli may be composed
of elements from different modalities. For example, an intermodal
compound might consist of a color and tone presented together. At
the highest  level of the
hierarchy are studies that use the dimensions
of space, time, and number (See Figure
1). These dimensions can be thought of as
more abstract than the dimensions at the intermodal and intramodal levels
because their signals can be carried by stimuli from any sensory modality
(Roberts, 1998). For instance, a spatial location can be indicated
by a visual stimulus presented at a specific location or by a tone emanating
from a specific location. Likewise, counting and timing can be carried
out with stimuli from either visual or auditory dimensions. These abstract dimensions may be combined with one another or with one
of the dimensions lower in the hierarchy to form compound stimuli. The
following section reviews findings of such experiments, as well as
the theoretical explanations proposed to account for the pigeons� performance.
level of the
hierarchy are studies that use the dimensions
of space, time, and number (See Figure
1). These dimensions can be thought of as
more abstract than the dimensions at the intermodal and intramodal levels
because their signals can be carried by stimuli from any sensory modality
(Roberts, 1998). For instance, a spatial location can be indicated
by a visual stimulus presented at a specific location or by a tone emanating
from a specific location. Likewise, counting and timing can be carried
out with stimuli from either visual or auditory dimensions. These abstract dimensions may be combined with one another or with one
of the dimensions lower in the hierarchy to form compound stimuli. The
following section reviews findings of such experiments, as well as
the theoretical explanations proposed to account for the pigeons� performance.
Intramodal
Studies
Most laboratory studies with pigeons have tested attention
to two visual elements using a delayed matching-to-sample procedure. In
delayed matching-to-sample, pigeons are trained to peck the one of two
comparison stimuli that is identical to a previously presented sample stimulus.
Pecking the correct comparison stimulus is followed by food. Suppose
a pigeon is placed in an operant chamber (Skinner box) that contains a
row of three circular keys along one wall. Any one of these keys
can be illuminated with a color or a pattern, as determined by a computer
program. For instance, the center key might be illuminated with red
light as the sample. After the sample is darkened (usually after
a certain amount of  sample exposure time or after a certain number of pecks
on the sample by the pigeon), two comparison stimuli are illuminated on
the side keys. One comparison might be colored red and the other
green. A peck to the red comparison (the comparison identical to
the sample) would result in the delivery of a food reward to the pigeon.
A peck to the green comparison would not be reinforced and would be followed
by darkness until the next trial began (see Grant & Kelly (2001)
for an
animated demonstration of delayed matching-to-sample).
sample exposure time or after a certain number of pecks
on the sample by the pigeon), two comparison stimuli are illuminated on
the side keys. One comparison might be colored red and the other
green. A peck to the red comparison (the comparison identical to
the sample) would result in the delivery of a food reward to the pigeon.
A peck to the green comparison would not be reinforced and would be followed
by darkness until the next trial began (see Grant & Kelly (2001)
for an
animated demonstration of delayed matching-to-sample).
In studies of attention, pigeons typically are trained on two different
element matching-to-sample problems and then are presented with samples
that are a combination of the two samples used in the training problems.
For example, after training on one delayed matching-to-sample problem with
red and blue keys as samples and comparisons and another delayed matching-to-sample
problem with vertical and horizontal lines as samples and comparisons,
possible compound samples could be vertical lines on a blue background,
horizontal lines on a red background, and so on. Memory for either
element of the compound (i.e. line orientation or color) can be tested
by presenting comparisons
from one of the  two dimensions (See Figure 2). For instance, a sample of
two dimensions (See Figure 2). For instance, a sample of horizontal
lines on a blue background could be followed by blue and red comparisons
(a color dimension test) or by vertical lines and horizontal lines on a
noncolored background as comparisons (a line orientation dimension test).
The correct choice would be either the blue comparison stimulus or the
horizontal lines stimulus. (To
experience an interactive demonstration of compound matching to sample click
here)
horizontal
lines on a blue background could be followed by blue and red comparisons
(a color dimension test) or by vertical lines and horizontal lines on a
noncolored background as comparisons (a line orientation dimension test).
The correct choice would be either the blue comparison stimulus or the
horizontal lines stimulus. (To
experience an interactive demonstration of compound matching to sample click
here)
Using similar line orientation and color stimuli, Maki and Leith (1973)
conducted such an intramodal test of compound stimulus processing.
They found that pigeons� matching accuracy was lower when the sample was
a compound than when it was one of the two elements (color or lines) used
in training. This decrement on compound sample trials was attributed
to a division of attention. The information processing capacity of
pigeons was assumed to be limited; Maki and Leith proposed that pigeons�
attention was divided between the color and line orientation elements of
the compound. This division of attention resulted in a weaker memory
for each element and therefore less accurate matching. A decrease
in performance on compound sample trials relative to element sample trials
became known as the divided attention effect. Could this detrimental
effect of divided attention on element matching performance be overcome?
Leith and Maki (1975) and Brown, Cook, Lamb, and Riley (1984) demonstrated
that sustained testing with one element of a compound increased performance
on the frequently tested element while performance on the non-tested element
suffered. Thus, pigeons could attend to one element of a compound,
if attention was biased toward that element, but otherwise performed worse
on compound tests relative to element tests.
Maki and Leith�s (1973) divided attention hypothesis soon was challenged
by others who suggested that the decrement on compound sample trials could
be explained on the basis of the pigeons� perception of the stimuli rather
than the amount of attention allocated to each element. Proponents
of a generalization decrement account of the divided attention effect argued
that it was the difference in appearance of the compound sample and the
element comparisons that caused pigeons to perform relatively poorly on
compound tests. Recall that in delayed matching-to-sample training problems,
the samples were identical to the comparisons, while the compound test
samples were not identical to the element comparisons that followed them.
Support for this view came from studies which varied the length of time
the sample was presented (Roberts & Grant, 1978; Santi, Grossi, &
Gibson, 1982). Recall that the divided attention hypothesis suggested
that subjects divided their attention between the two elements of a compound.
It should follow that increasing the amount of time the sample is illuminated
should allow a subject more time to process each element of the compound,
and matching performance should increase as the duration of the sample
increases. Contrary to this prediction, Roberts and Grant and Santi
et al. found that compound sample matching accuracy did not improve more
than element sample matching improved at longer sample durations.
These results were difficult for a divided attention account of the compound
sample matching decrement to explain and were used to support a generalization
decrement account.
While results from studies in which the duration of the sample was manipulated
weighed against the divided attention hypothesis, they did not directly
support a generalization decrement account. Grant and MacDonald (1986)
suggested a third hypothesis to explain the compound sample matching decrement.
Like the generalization decrement hypothesis, their interpretation was
based on the perceptual differences between compound stimuli and element
stimuli. Instead of the difference of interest being that between
the compound sample and the element comparisons (as was the focus of the
generalization decrement hypothesis), they focused on the difference between
the element samples used in training and the compound samples used in testing.
Grant and MacDonald�s account was based on the formation of prospective
codes during training. For example, in a matching-to-sample procedure,
a red key sample may come to elicit a prospective instruction or code to
peck a red comparison (see Grant & Kelly, this volume, for an
animated illustration of prospective coding). If a
horizontal line is then superimposed on the red key on test trials to form
a compound, the �peck red� code is only weakly elicited, and fewer correct
choices are made to a red comparison key. As a consequence, performance
on compound trials suffers. Additionally, a prospective coding account
predicts that differences in element and compound sample accuracy should  not be affected by sample duration: the same code should be elicited no
matter how long the sample is illuminated. In support of this account,
Grant and MacDonald found that pigeons trained with compound samples and
element comparisons showed a decrement in performance when tested with
element samples. Figure
3 illustrates
the training procedure used by Grant & MacDonald, and
Figure
4 shows the percentage of correct responses on element and compound
tests over blocks of test sessions. They accounted for this finding
by arguing that
not be affected by sample duration: the same code should be elicited no
matter how long the sample is illuminated. In support of this account,
Grant and MacDonald found that pigeons trained with compound samples and
element comparisons showed a decrement in performance when tested with
element samples. Figure
3 illustrates
the training procedure used by Grant & MacDonald, and
Figure
4 shows the percentage of correct responses on element and compound
tests over blocks of test sessions. They accounted for this finding
by arguing that the correct code was not elicited by element samples, even
though an element sample should not require the division of attention predicted
by the divided attention hypothesis. They also found that pigeons
showed the same decrement in performance even when the test sample remained
illuminated while the comparisons were presented (a simultaneous matching-to-sample
procedure). Allowing the sample to remain illuminated should give
pigeons plenty of time to process both dimensions of the compound.
According to the divided attention hypothesis, the result should be no
decrement in performance on compound test trials (an argument similar to
that tested by researchers testing the generalization decrement hypothesis
by lengthening the sample presentation time). If the sample simply
elicits a code to peck a comparison, however, a simultaneous procedure
should not eliminate the performance decrement. Grant and MacDonald
found that performance still declined on compound test trials. The coding
decrement hypothesis accounted for results that contradicted the divided
attention hypothesis.
the correct code was not elicited by element samples, even
though an element sample should not require the division of attention predicted
by the divided attention hypothesis. They also found that pigeons
showed the same decrement in performance even when the test sample remained
illuminated while the comparisons were presented (a simultaneous matching-to-sample
procedure). Allowing the sample to remain illuminated should give
pigeons plenty of time to process both dimensions of the compound.
According to the divided attention hypothesis, the result should be no
decrement in performance on compound test trials (an argument similar to
that tested by researchers testing the generalization decrement hypothesis
by lengthening the sample presentation time). If the sample simply
elicits a code to peck a comparison, however, a simultaneous procedure
should not eliminate the performance decrement. Grant and MacDonald
found that performance still declined on compound test trials. The coding
decrement hypothesis accounted for results that contradicted the divided
attention hypothesis.
In a more recent investigation of divided attention, Langley and Riley
(1993) have provided evidence for the original divided attention hypothesis
while controlling for generalization decrement and coding dec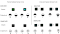 rement explanations.
They used a symbolic matching-to-sample procedure rather than the identity
matching procedure used in previous studies. The design of their experiment
can be seen in Figure
5. (To
experience an interactive demonstration of Langley and Riley's element and
compound tasks - click here). In one
task, blue or green key samples were followed by lit upper and lower halves
of comparison keys. In the other task, a triangle or circle sample
was followed by vertical and horizontal lines as comparisons. Since samples
and comparisons were taken from different dimensions, the comparisons should
be no more perceptually different from compound samples than from element
samples, thereby ruling out a stimulus generalization decrement explanation.
In order to rule out the possibility that pigeons were forming prospective
codes to element samples that would later fail to be elicited by compound
samples, subjects were trained with element and compound samples from the
beginning. With these procedural modifications, subjects still showed
poorer matching accuracy on compound stimulus trials than on trials with
both types of elements. It seems, therefore, that pigeons may indeed
be dividing their attention between the two elements of a compound stimulus.
This shared attention hypothesis and others suggested to account for the
compound sample test trial decrement are summarized in Table
1.
rement explanations.
They used a symbolic matching-to-sample procedure rather than the identity
matching procedure used in previous studies. The design of their experiment
can be seen in Figure
5. (To
experience an interactive demonstration of Langley and Riley's element and
compound tasks - click here). In one
task, blue or green key samples were followed by lit upper and lower halves
of comparison keys. In the other task, a triangle or circle sample
was followed by vertical and horizontal lines as comparisons. Since samples
and comparisons were taken from different dimensions, the comparisons should
be no more perceptually different from compound samples than from element
samples, thereby ruling out a stimulus generalization decrement explanation.
In order to rule out the possibility that pigeons were forming prospective
codes to element samples that would later fail to be elicited by compound
samples, subjects were trained with element and compound samples from the
beginning. With these procedural modifications, subjects still showed
poorer matching accuracy on compound stimulus trials than on trials with
both types of elements. It seems, therefore, that pigeons may indeed
be dividing their attention between the two elements of a compound stimulus.
This shared attention hypothesis and others suggested to account for the
compound sample test trial decrement are summarized in Table
1.
|
Table 1. Three hypotheses proposed to account for
pigeons'
accuracy deficits on compound sample matching tests |
|
Shared Attention
|
Maki & Leith (1973), Langley &
Riley (1993)
|
Attention is divided between the two elements
of a compound test sample, resulting in a weakening of memory for each
dimension and therefore less accurate matching.
|
|
Generalization Decrement
|
Roberts & Grant (1978), Santi
et al (1982)
|
On compound test trials, the difference
in appearance between the compound sample and the element comparisons results
in less accurate matching relative to element sample trials.
|
|
Coding Decrement
|
Grant & MacDonald (1986)
|
Compound samples do not elicit the same
prospective code as do element samples resulting in less accurate matching
on compound sample trials relative to element sample trials.
|
Recently, Roberts (1998) has offered an interpretation of the
shared attention hypothesis that may account for seemingly contradictory
findings, especially those from experiments in which the duration of the
sample was lengthened. He suggested that switching back and forth
between processing one dimension and another dimension may results in decay
of the memory for the non-processed dimension at any given point during
a sample presentation. If repeated switches in processing continued
for the entire duration of the sample, the deficit in accuracy on compound
trials relative to element trials should persist at longer sample durations.
Also, presenting the sample and comparison stimuli simultaneously should
not necessarily produce higher accuracy with compound samples, since the
animal would still switch back and forth between the two elements of the
compound, resulting in lower accuracy on each one.
Intermodal
Studies
An intermodal compound stimulus is made up of two elements from different
sensory modalities. For example, Kraemer and Roberts (1985) conducted
an intermodal study by combining stimuli from the visual and auditory modalities.
A group of pigeons was trained on two matching-to-sample procedures with
sample stimuli that consisted of the color of a key (red or green) in one
task and the frequency of a tone (high or low) in the other. Notice
that this experiment again used a symbolic matching-to-sample procedure
in which the samples and comparisons were not identical. Thus, pigeons
had to choose between vertical and horizontal line comparison stimuli following
red and green sample stimuli and between yellow and blue comparison stimuli
following high and low tones. When the tone and color samples were
presented in compound and only one dimension was tested, pigeons� accuracy
did not decline on color tests, but performance was significantly worse
on tone frequency tests than performance on tone alone control tests.
Unlike previous studies that used two visual samples to make up a compound,
Kraemer and Roberts found that one dimension (color) seemed to dominate
processing of a tone/color compound. Interestingly, Foree and LoLordo
(1973) and Shapiro, Jacobs, and LoLordo (1980) found that tone stimuli
acquired more control over responding than did light when the reinforcer
was shock avoidance. These findings illustrate that the stimulus
that controls responding can differ depending on the reinforcer.
Studies
with Abstract Dimensions
As illustrated in the hierarchical framework, the dimensions
of space, time, and number may also be used as elements in compound stimuli.
Kraemer, Mazmanian, and Roberts (1987) tested pigeons with compound stimuli
and used the abstract dimension of space as one element. Pigeons
were trained with left and right keys illuminated with white light as samples
and a triangle pattern and three dots in a diagonal pattern as comparisons.
Pigeons matched based on the left-right location of the sample; thus, the
triangle would be the correct comparison choice after the sample appeared
on the right key, but the diagonal dot pattern would be correct after the
sample appeared on the left key. In another task, the birds were
trained to match red and green comparisons to blue or yellow colored samples
on the center key. Here, they matched based on the color of the sample.
After learning both tasks, pigeons were tested with compound samples consisting
of blue or yellow illumination on the left or right key. Pigeons
showed no deficit in matching accuracy on test trials in which the dimension
tested was either color or space. Spatial location and color appear
to have been processed simultaneously, with no decrement on either dimension.
Sutton and Roberts (1998) investigated attention to compound stimuli
composed of time and visual pattern elements (Experiments 1 and 3a) and
time and spatial location elements (Experiments 2 and 3b). They found
that pigeons could process the duration of a stimulus and the orientation
of a line (vertical or horizontal) presented on that stimulus simultaneously.
The subjects required explicit duration training with the line stimulus,
however, in order to accomplish this simultaneous processing. That
is, they were trained on a duration matching problem with a white center
key illuminated for 2 or 10 seconds as the sample and with red and green
comparison keys. The same pigeons also were trained on a line orientation
matching problem, with a vertical or horizontal line on the center key
as the sample and with choice between yellow and blue comparison stimuli. On compound tests, the vertical or horizontal line was presented for 2
or 10 seconds, and either the line orientation dimension was tested by
presenting yellow and blue comparisons, or the time duration dimension
was tested by presenting red and green comparison stimuli. The pigeons
were unable to accurately match the duration of these compounds.
It was felt that pigeons might need temporal training with the specific
visual stimuli used in the compounds. Thus, these pigeons then were
given further training to match the duration of a vertical or horizontal
line stimulus. Eventually they were tested on time/line orientation
compounds in sessions where they were unaware of the dimension tested on
a given trial until the comparisons were illuminated. Now, pigeons
matched the time element of the samples as accurately as they matched time
in sessions that contained only time tests. The same pattern of results
was found with birds tested on space/time compounds; birds needed explicit
duration training with left and right  stimuli in order to process both
dimensions simultaneously. Results of these tests are presented in Figure
6. Table 2 summarizes the
test procedures used by Sutton and Roberts (1998). The results of
tests with both compounds suggest that attention needs to be directed to
the time dimension of a stimulus in order to match based on duration, but
that simultaneous processing is easily shown once that training has occurred.
stimuli in order to process both
dimensions simultaneously. Results of these tests are presented in Figure
6. Table 2 summarizes the
test procedures used by Sutton and Roberts (1998). The results of
tests with both compounds suggest that attention needs to be directed to
the time dimension of a stimulus in order to match based on duration, but
that simultaneous processing is easily shown once that training has occurred.
|
Table 2. Sutton & Roberts
(1998) test procedure (Experiments 3a and 3b) |
| |
Line orientation/duration test |
Spatial location/duration test |
| Sample |
Vertical or Horizontal for 2 or 10 sec |
Left or Right white stimulus for 2 or
10 sec |
| Comparisons |
Duration test (red and green) or
Line orientation test (blue and yellow) |
Duration test (red and green) or
Spatial location test (blue and yellow) |
IV. Other
Studies That Combine Abstract Dimensions Within The Hierarchical Framework
A few other studies with pigeons have combined abstract
dimensions while not directly focusing on attention to each dimension.
The intention often was to determine simply whether an animal attended
to both dimensions presented or how it might average information from different
dimensional cues presented simultaneously. For
instance, Cheng, Spetch, and Miceli (1996) used a touch screen apparatus
to present a compound stimulus using the dimensions of time and space.
On training trials, a rectangle moved across the screen at a constant rate
of 1 cm per second. Subjects were rewarded for the first peck to
the target after 10 seconds had passed. Since the rectangle was moving
at a constant rate, not only could the pigeons use how much time had passed
to determine when to respond, but they could also attend to the location
of the rectangle, since it was always in the same location just prior to
reinforcement. Cheng et al. were interested in which cue, if not
both, the pigeons used to determine when to peck. On test trials,
the rectangle moved across the screen faster or slower than in training.
Specific predictions about the peak of the pigeons� responding were made
based on whether they were using time or location. That is, if the
subjects were using the duration of the stimulus presentation, the response
peak on slow rate tests should have been before the target had passed its
location of reinforcement from training. On fast rate tests, the
peak should occur after the rectangle had reached the same location.
Alternatively, if the pigeons were using the location of the stimulus to
determine when to peck, their responding should peak late (in terms of
time) on slow trials and early on fast trials. The results showed
that the pigeons tended to average the two dimensions but showed a slight
bias toward the time cue. Cheng et al. concluded that the pigeons
had learned to use both the time and space cues in training to determine
when to respond.
Time and number were investigated in compound by Roberts and Mitchell
(1994) with pigeons. Pigeons were trained with flashes of a houselight
that flashed at the rate of 1/second. Pecking on one key was reinforced
after two flashes in 2 seconds and pecking on another key was reinforced
after eight flashes in 8 seconds. Thus, time and number cues were
confounded, in that the low number and short time cue signaled reward
for one response and the large number and long time cue signaled reward
for the other response. To unconfound the time and number dimensions,
tests were performed in which one dimension was held constant, while the
other dimension was varied. To measure control by time, tests consisted
of four flashes that lasted for 2, 3, 4, 5, 6, 7, or 8 seconds. To
measure control by number, tests consisted of a 4-second presentation of
2, 3, 4, 5, 6, 7, or 8 flashes. Psychophysical curves plotted proportion
of trials on which the large number/long time key was chosen as a function
of time and number. These curves showed equivalent control by time
and number. Meck and Church (1983) found similar results with rats,
while Breukelaar and Dalrymple-Alford (1998) found that rats� attention
to time may be stronger than attention to number.
V.
Future
Directions
While the abstract dimensions of time, space, and number have been investigated
as elements in compound sample stimuli, only a few studies have directly
focused on attention to these dimensions. The few studies reviewed
here suggest that attentional processes may interact in a complex fashion
with the types of dimensions placed in compound with one another.
Future research on the processing of multiple dimensions and the limits
to that processing may further demonstrate how pigeons process different
types of information simultaneously and may provide more clues to the organization
of information processing systems. Pigeons� attention is most certainly
divided between widely varying stimulus types in nature, and survival may
depend on the appropriate division of attentional resources. Future
studies may also address the limits to divisions of attention in nature
as well as in the laboratory.
VI.
References
Bitterman, M. E., & Coate, W. B. (1950).
Some new experiments on the nature of discrimination learning in the rat.
Journal
of Comparative and Physiological Psychology, 43, 198-210.
Cheng, K., Spetch, M. L., & Miceli, P. (1996). Averaging
temporal duration and spatial position. Journal of Experimental
Psychology: Animal Behavior Processes, 22, 175-182.
Cherry, E. C. (1953). Some experiments on the recognition of speech
with one and with two ears. Journal of the Acoustical Society of America,
25, 275-279.
Dukas, R. (1998). Constraints on information processing and their
effects on behavior. In R. Dukas (Ed.), Cognitive Ecology: The evolutionary
ecology of information processing and decision making (pp.
89-127). Chicago, IL: University of Chicago Press.
Ehrenfreund, D. (1948). An experimental test of the continuity
theory of discrimination learning with pattern vision. Journal
of Comparative and Physiological Psychology, 41, 408-422.
Foree, D. D., & LoLordo, V. M. (1973). Attention
in the pigeon: The differential effects of food-getting vs. shock avoidance
procedures. Journal of Comparative and Physiological Psychology,
85, 551-558.
Grant, D. S., & MacDonald, S. E. (1986). Matching to element and
compound samples in pigeons: The roles of sample coding. Journal of
Experimental Psychology: Animal Behavior Processes, 12, 160-171.
Kraemer, P. J., Mazmanian, D. S., & Roberts, W. A. (1987). Simultaneous
processing of visual and spatial stimuli in pigeons. Animal Learning
& Behavior, 15, 417-422.
Kraemer, P. J., & Roberts, W. A. (1985). Short-term
memory for simultaneously presented visual and auditory signals in the
pigeon. Journal of Experimental Psychology: Animal Behavior Processes,
11, 137-152.
Krechevsky, I. (1938). A study of the continuity of the problem-solving
process. Psychological Review, 45, 107-133.
Langley, C. M., & Riley, D. A. (1993). Limited capacity information
processing and pigeon matching-to-sample: Testing alternative hypotheses.
Animal
Learning & Behavior, 21, 226-232.
Leith, C. R., & Maki, W. S. (1975). Attention shifts during
matching-to-sample performance in pigeons. Animal Learning &
Behavior, 3, 85-89.
Lindsay, P. H., Taylor, M. M., & Forbes, S. M. (1968). Attention
and multidimensional discrimination. Perception and Psychophysics,
4, 113-117.
Maki, W. S., & Leith, C. R. (1973). Shared
attention in pigeons.
Journal of the Experimental Analysis of Behavior,
19, 345-349.
Meck, W. H., & Church, R. M. (1983). A
mode control model of counting and timing processes. Journal of
Experimental Processes: Animal Behavior Processes, 9, 320-334.
Riley, D. A., & Leith, C. R. (1976). Multidimensional psychophysics
and selective attention in animals. Psychological Bulletin, 83,
138-160.
Roberts, W. A. (1998). Priciples of animal cognition.
Boston, MA:
McGraw-Hill.
Roberts, W. A., & Grant, D. S. (1978). Interaction of sample and
comparison stimuli in delayed matching-to-sample with pigeons. Journal
of Experimental Psychology: Animal Behavior Processes, 4, 68-82.
Roberts, W. A., & Mitchell, S. (1994). Can a pigeon simultaneously
process temporal and numerical information? Journal of Experimental
Psychology: Animal Behavior Processes, 20, 66-78.
Santi, A., Grossi, V., & Gibson, M. (1982). Differences in matching-to-sample
performance with element and compound sample stimuli in pigeons. Learning
and Motivation, 13, 240-256.
Shapiro, K. L., Jacobs, W. J., & LoLordo, V. M. (1980). Stimulus-reinforcer
interactions in Pavlovian conditioning of pigeons: Implications for selective
associations. Animal Learning & Behavior, 8, 586-594.
Spence, K.W. (1936). The nature of discrimination
learning in animals. Psychological Review, 43, 427-449.
Spence, K. W. (1945). An experimental test of the continuity and
non-continuity theories of discrimination learning. Journal of
Experimental Psychology, 35, 253-266.
Sutherland, N. S., & Mackintosh, N. J. (1971). Mechanisms of
animal discrimination learning. San Diego, CA: Academic Press.
Sutton, J. E. & Roberts, W. A. (1998). Do pigeons show incidental
timing? Some experiments and a suggested hierarchical framework for the
study of attention in animal cognition. Behavioral Processes, 44, 263-275.
Treisman, A. (1969). Strategies and models of selective
attention.
Psychological
Review, 76, 282-299.
|
Acknowledgement
Preparation of this chapter was supported by an operating grant
from the Natural Sciences and Engineering Research Council of Canada
to W. A. Roberts. We thank Bob Cook for his assistance with the
interactive demonstrations and formatting of this chapter.
|